一 : 微生物阿魏酸酯酶的研究进展
专题论述
中国酿造
2008年第10期
总第187期
?11 ?
微生物阿魏酸酯酶的研究进展
许 晖,孙兰萍,张 斌,石亚中
(蚌埠学院食品与生物工程系,安徽蚌埠,233030)
摘 要:阿魏酸酯酶能水解阿魏酸甲酯、低聚糖阿魏酸酯和多糖阿魏酸酯中的酯键,将阿魏酸游离出来的一种酶,在农业、食品和制药工业等具有广阔的应用前景。[www.61k.com)该文介绍了阿魏酸酯酶的微生物来源和生产、理化性质、结构特征、酶活力测定等方面的国内外研究进展。关 键 词:阿魏酸酯酶;微生物发酵法;研究进展
中图分类号:Q55 文献标识码:A 文章编号:0254-5071(2008)10-0011-05
Advanceresearchonferuloylesterasesproducedbymicrobes
XUHui,SUNLanping,ZHANGBin,SHIYazhong
(DepartmentofFoodandBioengineering,BengbuCollege,Bengbu233030,China)
Abstract:Ferulicacidesterases(FAE)isanewenzyme,whichcanhydrolyzemethylferulate,oligosaccharidesferulateandpolysaccharidesferulatetoferulicacid(FA),andthereforeplayamajorroleinthedegradationofplantbiomass1Italsoshowsperspectiveapplicationinagriculture,foodandpharmaceuticalindustries1Thispaperreviewedtheresearchprogressandtrendinrecentstudiesrelatedtomainenzyme2producingmicroorganisms,physicochemicalproperties,molecularstructure,catalysischaracteristics,productiontechniquesandassayofmicrobialFAE1Keywords:ferulicacidesterase;microbialfermentationmethod;researchadvance
阿魏酸(ferulicacid)的化学名称为42羟基232甲基222苯丙烯酸,是植物界普遍存在的一种酚酸,构中起着重要的作用,素之间、形成交接,硬。在谷物的细胞壁中,以酯键的形式连接在阿拉伯木聚糖的阿拉伯糖残基上[122]。其主要的生理功能有抗氧化和清除自由基、抗血栓、降血脂及防治冠心病,并具有抗菌消炎以及抗突变和防癌作用[3]。
,目前在医生产阿魏酸的方法有碱法和酶法,其中通过酶解法生产阿魏酸受到许多学者的重视。
阿魏酸酯酶(E1C131111173,ferulicacidesterase,FAE)又称肉桂酸酯酶,是最近才分离纯化出来的一种酯酶,它是羧酸酯水解酶的一个亚类,也是一种胞外酶,是能水解阿魏酸甲酯、低聚糖阿魏酸酯和多糖阿魏酸酯中的酯键,将阿魏酸游离出来的酶[425]。在食品工业中可利用阿魏
收稿日期:2007211201
基金项目:蚌埠学院微生物学重点课程建设项目,蚌埠学院中青年学科带头人培养对象专项资助(院人字[2006]18号)作者简介:许 晖(19692),男,安徽蚌埠人,副教授,主要从事食品科学及食品生物技术研究工作。
[30]刘振扬,高洪平,刘文俊,等1啤酒废酵母酶法生产酱油技术[J]1酿酒科东北大学学报:自然科学版,2004,25(1):892911
[40]邱延省,成先雄1啤酒酵母吸附镉离子的实验研究[J]1环境污染与防
技,2006(11):972981
[31]李 魁1利用燕麦和啤酒废弃生物质酿造酱油的研究[J]1中国酿造,
2006(2):372391
[32]陈 云1啤酒废酵母在酱油生产中的应用[J]1江苏调味副食品,2005,
22(2):22223,311
[33]刘 超,刘振扬,吴润姣,等1啤酒废酵母酶法制取酵母浸膏技术[J]1酿酒
治,2004,26(2):952971
[41]徐惠娟,廖生赟,龙敏南,等1啤酒酵母吸附镉的研究[J]1工业微生物,
2004,34(2):102141
[42]刘振扬,刘 超1啤酒酵母在处理含镉工业废水中的应用研究[J]1酿酒
科技,2006(7):862881
[43]王战勇,张 晶,苏婷婷1啤酒废酵母对Pb2+的吸附[J]1辽宁石油化工
科技,2006(12):10421051
[34]李大鹏1啤酒酵母营养饮料的研制[J]1粮食与食品工业,2005,12(2):
24226,311
[35]李 彦,李巍青,俞 健,等1用啤酒废酵母泥生产鲜味食用蛋白的生产
大学学报,2006,26(1):262291
[44]王战勇,张 晶,苏婷婷1啤酒废酵母对铜离子的吸附研究[J]1南昌大
学学报:工科版,2007,29(1):622651
[45]夏良树,陈仲清1啤酒酵母菌—活性污泥曝气工艺处理含铀废水研究[J]
工艺[J]1食品研究与开发,2004,25(5):10621081
[36]李 彦1利用啤酒废酵母泥生产营养酸奶[J]1包装与食品机械,2004,
22(5):332351
[37]孙凌雪1啤酒废酵母生产酵母蛋白营养粉的工艺研究[J]1酿酒,2004,
31(5):962981
[38]张秀丽,刘月英1贵、重金属的生物吸附[J]1应用与环境生物学报,
2002,8(6):66826711
)的生物吸附[J]1[39]周东琴,朱一民,魏德州,等1啤酒酵母菌对汞离子(Ⅱ
1核技术,2006,29(11):87228761
[46]张志强,林 波1利用啤酒废水所产微生物絮凝剂处理靛蓝印染废水的
研究[J]1环境污染与防治,2006,28(2):14921511
[47]郑爱榕,赵立清,詹力扬,等1利用啤酒废水养殖螺旋藻研究[J]1海洋科
学,2004,28(7):262311
[48]王江波1麒麟啤酒化腐朽为神奇开发环保燃料[J]1食品科技,2007,32
(2):198

1
阿魏是什么 微生物阿魏酸酯酶的研究进展
?12 ?
2008No110
SerialNo1187
ChinaBrewing
SpecialDiscuss
酸酯酶打断阿魏酸与细胞壁材料如麸皮、秸秆中多糖的交联,高效降解多糖并获得反式阿魏酸,它和阿拉伯木聚糖酶联合作用的水解产物低聚糖(主要是低聚木糖)、阿魏酸都是重要的功能性食品基料,戊糖可用于酒精发酵、生产木糖醇等。(www.61k.com]在饲料工业中,利用阿魏酸酯酶处理植物性的原材料,可以将阿魏酸从植物细胞壁的结构中游离出来,从而破坏细胞壁的骨架结构,使得木质素、半纤维素及纤维素之间的连接被部分打破,结构变得比处理前疏松,变得疏松的原料更容易被牲畜消化吸收利用,可以提高饲料的利用效率。本文就阿魏酸酯酶的微生物来源和生产、理化性质、结构特征、催化机理、酶活力测定等方面进行了综述。1阿魏酸酯酶的微生物来源
1987年MACKENZIECR等[6]培养橄榄色链霉菌
(Streptomycesolivochromogenes)时,发现阿魏酸酯酶可
61阅读提醒您本文地址:
以从麦麸中释放出阿魏酸,此后阿魏酸酯酶被认为是半纤维素水解酶。研究表明真菌、细菌和酵母都能分泌阿魏酸酯酶,目前发现的产酶微生物主要有黑曲霉(Aaspergillusniger)、链霉菌(如Streptomycesavermiti2lis)、梭菌(如Clostridiumthermocellum)、杆菌(如Bacil2lussp1)、乳酸杆菌(Lactobacilli)、假单胞菌(如Pseudo2monasfluorescens)等。但绝大多数阿魏酸酯酶从真菌中分离得到,尤其曲霉菌属(Aspergillussp1),如黄柄曲霉(Aspergillusflavipes)和黑曲霉(Aspergillusniger),特别受到研究者的普遍关注,这2种菌种能以去淀粉麦麸或玉米糠为碳源,通过深层培养产生阿魏酸酯酶[728]。阿魏酸酯酶微生物来源以及培养条件见表1。
表1产阿魏酸酯酶微生物来源[9]
Table11ProductionofFAEsbyxylanolyticmicroorganisms
产酶微生物
A1awamoriVTT2D271025A1flavipes
AspergillusfoetidusVTT2D271002A1nigerA1niger
A1nigerCBS120149A1nigerI21472A1nigerI21472
AspergillusoryzaeVTT2D285248
AspergillusphoenicisIMI211395AspergillusterreusVTT2D282209A1pullulansNRRLY231121
碳源
3%麦麸1%麦麸3%纤维素粉
30℃、7d287d
/(mU?mL21)
68201828415340663007240002434715113150109833146191671117552201201542914801614560411216180144801200210156273000
方法
SmFSmFSmFSmFSSFSmFSmFSSFSmFSmFSmFSmFSmFSmFSmFSmFSmFSmFSmFSmFSmFSmFSmFSSFSSFSSFSSFSmFSmFSmFSmFSmFSmFSmFSSFSmFSmF
μ20g/mL75g)
%+1%5%30%甜菜渣3%麦麸
1%MFG+015%葡萄糖3%纤维素粉1%桦木木聚糖015%桦木木聚糖2%玉米芯1%玉米皮1%玉米皮
015%葡萄糖+012%乳酸0105%阿魏酸甲酯2%纤维素
014%绊根草汁+014%剑麻浆1%甜菜渣20%啤酒糟(215g)33%麦麸(75g)
33%(燕麦麸+麦麸,7∶3)(150g)33%麦麸(75g)
014%绊根草汁+014%剑麻浆1%去淀粉麦麸1%去淀粉麦麸1%甘蔗渣1%去淀粉麦麸1%去淀粉麦麸115%去淀粉麦麸22%(500g)麦秸杆2%甜菜渣
1%酥油草汁+015%葡萄糖
、4d
26℃、7d、酶提取液400mL100mL、25℃、3d200mL、30℃、3d
30℃、4d、酶提取液30mL/5g物料30℃、7d50mL、30℃、7d30℃、7d2L、30℃、60h
100mL、30℃、24h、CFE16L、65℃
300mLor10L、39℃、50h17L、30℃、70h100mL、30℃、5d100mL、30℃、5d1L、37℃、60h
100mL、30℃、24h、CFE1175L、39℃、5d39℃、5d2L、25℃、6d
30℃、196h、提取液10mL/1g物料26℃、7d、提取液400mL30℃、12d、提取液300mL26℃、7d、提取液400mL39℃、5d1L、30℃、14d30mL、37℃、1d?2d30mL、37℃、4d1L、37℃、3d1L、37℃、3d20mL、30℃、4d
50℃、163h、提取液10mL/

1g物料25℃、7d50mL、30℃、7d
BacillussubstilisATCC76610105%阿魏酸甲酯
Clostridiumstercorarium
NCIMB11754
FibrobactersuccinogenesS851F1oxysporumF3
F1proliferatumNRRL26517FusariumverticillioidesNRRL26517
014%微晶纤维素粉
LactobacillusacidophilusIFO13951LactobacillusfermentumNCFB1751NeocallimastixMC22OrpinomycesPC21
PenicilliumfuniculosumIMI2134756PenicilliumbrasilianumIBT20888PenicilliumbrevicompactumPenicilliumpinophilumCMI87160P1expansumPiromycesMC21S1communeATCC38548
StreptomycesavermitilisCECT3339S1avermitilisUAH30StreptomycesC2248
S1olivochromogenesNRCC2258Streptomycessp1S10S1thermophileATCC34628T1stipitatusCBS375148TrichodermareeseiQM9414
阿魏是什么 微生物阿魏酸酯酶的研究进展
专题论述
中国酿造
2008年第10期
总第187期
表2部分微生物产阿魏酸酯酶的酶学特性Table21EnzymaticcharacteristicsofpurifiedFAEs
?13 ?
在酶制剂生产的各种方法中,固态发酵技术(Solid
statefermentation,SSF)引起许多学者重视的,特别是真菌的培养,因为SSF具有生产效率高、工艺简单、操作粗放、能耗少、废液少、产物分离较容易等优点。(www.61k.com)固态发酵最重要的是选择合适的基质,用于阿魏酸酯酶生产的固态基质主要是含淀粉基和纤维素基的农产品或农业资源,如甜菜渣、麦麸、麦秸杆和啤酒糟,这些基质不仅作为载体,还作为一些微生物生长需要的营养物质,在这些固态基质培养产生阿魏酸酯酶的微生物有A1niger[10211]、青霉菌属(Penicilli2
[10,12214]um)和侧孢霉属(Sporotrichum)[15],其中Penicilliumbrasilianum在固态培养阿魏酸酯酶的产量最大。
培养基质的选择对微生物是否能够产生阿魏酸酯酶至关重要。单糖和二糖(如葡萄糖、木糖、乳糖、麦芽糖和木醇糖)作为基质时,微生物不能产生阿魏酸酯酶,这可能是由于葡萄糖分解代谢物的阻遏作用。含有大量酯化阿魏酸的混合碳源,如麦麸[16217]、去淀粉麦麸[18219]、玉米皮[20221]、啤酒糟[12,22]和甜菜渣[17,21]等,最适合用作基质生产阿魏酸酯酶。影响,生物合成。尖孢镰孢()养时,(de2esterifiedcorn2cobs)的1/515;若把游离的阿魏酸添加到脱酯玉米芯中,则阿魏酸酯酶的产量提高了115倍,结果表明阿魏酸的存在对F1oxysporum产生阿魏酸酯酶影响不显著,但是如果从培养基质脱除阿魏酸,会造成阿魏酸酯酶产量的降低。总之,阿魏酸酯键可能是诱导阿魏酸酯酶产生的主要因素,如果以纯化的木聚糖,如燕麦木聚糖(oatspeltxylan)[22]、桦木木聚糖(birchwoodxylan)[23]和落叶松木聚糖(larchwoodxy2lan)[24]作为培养基质时,阿魏酸酯酶的产量明显降低。目前纤维素也可用于产生阿魏酸酯酶[25226]。2阿魏酸酯酶的理化性质
1991年,FAULDSCB等[27]第1次报道了分离纯化的橄榄色链霉菌(Streptomycesolivochromogenes)阿魏酸酯酶,至今约有30种微生物产阿魏酸酯酶被分离纯化,如泡盛曲霉(Aspergillusawamori)、黑曲霉(Aspergillusniger)、构巢曲霉(Aspergillusnidulans)、米曲霉(Aspergillusoryzae)、塔宾曲霉(Aspergillustubingensis)、出芽短梗霉(Aureobasidiumpullulans)、耐热梭状芽孢杆菌(Clostridi2umstercorarium)、热纤梭菌(Clostridiumthermocellum)、尖孢镰孢(Fusariumoxysporum)、层出镰刀菌(Fusariumpro2liferatum)、嗜酸乳杆菌(Lactobacillusacidophilus)、绳状青霉(Penicilliumfuniculosum)、真菌嗜松青霉(Penicilliumpinophilum)、厌氧真菌(Piromycesequi)、新美丽丝菌(Neo2callimastixMC22)、粗糙脉孢菌(Neurosporacrassa)和Ta2laromycesstipitatus。部分微生物产阿魏酸酯酶的酶学特性,见表2。
61阅读提醒您本文地址:
产酶微生物
A1nigerA1pullulansF1oxysporumP1pinophilumS1thermophileS1olivochromogenes
相对分子质量/kDa
3621031573329等电点
313615915416315719最适pH值
510617710610610515最适温度/
℃
5560555550~6030不同微生物来源阿魏酸酯酶性质不同,一般微生物产阿魏酸酯酶的相对分子质量为30kDa~70kDa,但是
A1pullulans所产阿魏酸酯酶相对分子质量为210kDa,NeocallimastixMC22所产阿魏酸酯酶相对分子质量为
11kDa。阿魏酸酯酶的最适作用pH值为410~810,最适作
用温度为30℃~65℃。
33],热纤梭菌(Clostridium
)是一种嗜热的厌氧细菌,能产生一种叫纤维
小体(cellulosome)的多酶复合物,该多酶复合物不仅具有纤维素水解活性,还具有木聚糖酶、甘露聚糖酶、几丁质酶、果胶裂解酶和β2葡萄糖苷酶活力[29],BLUMDL等[30]证明了热纤梭菌纤维小体还具有阿魏酸酯酶的活力,木聚糖酶10A(Xyn10A)和木聚糖酶10B(Xyn10B)是组成阿魏酸酯酶的主要部分。热纤梭菌产木聚糖酶具有2种不同作用的酶,包含2种不同的催化机制:一种是水解木聚糖主链的糖苷键;另一种是水解连接阿拉伯糖苷侧链和阿魏酸的酯键。Prates等[31]对阿魏酸酯酶结构进行了研究,由于其催化底
物的结构复杂,相应的阿魏酸酯酶也有着相当复杂的结构,虽然不同的阿魏酸酯酶结构有差异,但都为球蛋白质,且大都具有Ser2His2Asp的“三点催化活性中心”,阿魏酸酯酶Xyn10B的三维结构见图1
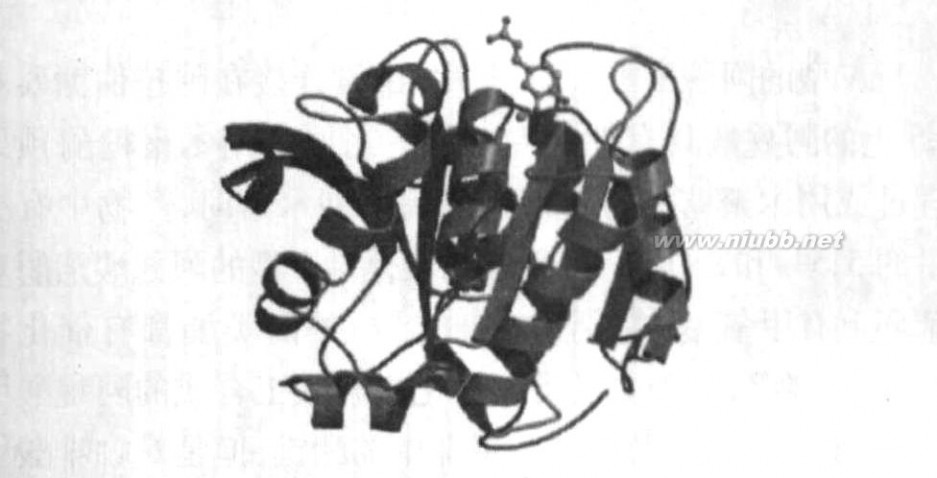
。
图1阿魏酸酯酶的三维结构
Fig11Three2dimensionalstructureofFAE
在催化过程中,阿魏酸酯酶的催化中心和辅助中心将阿魏酸酯化合物中的阿魏酸残基束缚,而后经过一系列化学过程将酯键断裂,使阿魏酸游离出来。阿魏酸酯酶催化机理见图2[31]

。
阿魏是什么 微生物阿魏酸酯酶的研究进展
?14 ?
2008No110
SerialNo1187
ChinaBrewing
Special

Discuss
型阿魏酸酯酶对1,52阿魏酸阿拉伯糖酯键也有催化作用,但效果远远低于A型。(www.61k.com)另外B型的阿魏酸酯酶不能使阿魏酸二聚体游离出来。B型阿魏酸酯酶还可以催化水解人工合成的阿魏酸甲酯、咖啡酸甲酯以及香兰酸甲酯中的酯键,但是对芥子酸甲酯无效。B型阿魏酸酯酶的产酶微生物主要有黑曲霉、青霉和链孢霉菌等。31213第3种类型
目前还发现一种称作XYLD的乙酸酯酶具有阿魏酸酯酶活性,它是从荧光假单胞杆菌的一个亚种中分离得到的,对苯乙烯酚酸酯类(阿魏酸酯、咖啡酸酯、香兰酸酯以及芥子酸酯)的化合物同样有催化作用。用XYLD处理小麦的外壳,产物中有5,5’2阿魏酸二聚体。
此外,也有研究者依据诱导产酶培养基、能水解的酯键、肽链中氨基酸的排列和产物中是否产生阿魏酸的二聚A、B、C和D4类[9]。4(U)是指在温度
μg,水解去淀粉玉米麸皮1min产生1低聚糖阿魏酸酯和去淀粉麦麸等天
然基质常用于测定阿魏酸酯酶活力的底物,人工合成酯类物质(如阿魏酸甲酯和阿魏酸乙酯)也用于酶活力测定的底物。阿魏酸含量测定方法以高效液相色谱法最为普遍,其他还有气相色谱法、薄层扫描法、分光光度法和高效毛细管电泳法等[24,33235]。
5阿魏酸酯酶在食品工业中的应用
图2阿魏酸酯酶的催化机理
Fig21CatalyticmechanismofFAE
312阿魏酸酯酶的分类和催化专一性
阿魏酸酯酶对苯乙酸酯类具有催化专一性,然而不同的阿魏酸酯酶的催化专一性也有所不同,依据催化专一性的不同,阿魏酸酯酶最初被分为A、B2种类型1后来又发现了一些其他类型[32]。31211A型
A型的阿魏酸酯酶对以1,52酯键连接在阿拉伯糖残基C5上的阿魏酸具有催化专一性。当原料用木聚糖酶预处理过或用木聚糖酶与阿魏酸酯酶协同水解时,产物中有少量的5,5’2和82O24’2阿魏酸二聚体。A型的阿魏酸酯酶对苯环上有甲氧基(特别是在3,5位)的酚类酯都有催化特性。A型阿魏酸酯酶还可以催化水解人工合成的阿魏酸甲酯、芥子酸甲酯以及香兰酸甲酯中的酯键,但是对咖啡酸甲酯无效。当以谷物作为培养基时,微生物倾向于产生A型阿魏酸酯酶。31212B型
以甜菜渣做培养基时,微生物倾向于分泌B型阿魏酸酯酶。B型的阿魏酸酯酶对连接在阿拉伯糖残基C2和半乳糖残基C6上的阿魏酸酯键具有专一性,黑曲霉来源的B
阿魏酸是天然抗氧化剂,也是近年来国际认知的防癌物质。其具有很好的抗氧化活性,对过氧化氢、超氧自由基、羟自由基、过氧化亚硝基都有强烈的清除作用,且能调节生理机能,抑制产生自由基的酶,增加清除自由基酶的活性。同时,阿魏酸可以作为合成天然香兰素的前体,以阿魏酸为原料,利用微生物的方法生产的香兰素与合成的香兰素相比,具有毒性低,安全性高的特点,可以用作食品和化妆品行业的香料[32,36]。
阿魏酸广泛存在于食品原料中,目前已经使用阿魏酸酯酶或与其他酶协同作用从各种农作物的副产品中如麦麸、玉米麸皮、啤酒糟、谷物纤维、甜菜渣、燕麦壳和苹果渣等作为原材料来提取阿魏酸[9,37238]。6阿魏酸酯酶的研究现状和展望
国外在阿魏酸酯酶的研究上已取得了一些卓有成效的工作。首先筛选了一批产阿魏酸酯酶的微生物;其次对部分微生物产阿魏酸酯酶进行了纯化,并对其酶学性质进行了研究,如酶的结构、最适温度、最适pH值及影响酶稳定性的其他因素;第3探讨了阿魏酸酯酶与一些多糖降解酶的协同作用;第4探讨了微生物产酶的影响因素和酶的工业化分离方法。
对阿魏酸酯酶的研究仍有很多工作要做。

首先要筛选
阿魏是什么 微生物阿魏酸酯酶的研究进展
专题论述
中国酿造
61阅读提醒您本文地址:
2008年第10期
总第187期
?15 ?
出产阿魏酸酯酶较高的微生物,因为目前大多数微生物发酵
产生阿魏酸酯酶的量仍然达不到工业化生产的要求,利用现代生物技术来改良微生物,可以提高阿魏酸酯酶的产量,目前已有研究者定位了与阿魏酸酯酶合成有关的基因,为进一步的研究奠定了基础;其次目前阿魏酸酯酶的应用还集中在食品工业中,在其他行业上应用的研究还有待开展。(www.61k.com)参考文献:
[1]RONALDP,VRIESD,VISSERJ1AspergilusEnzymesInvolvedinDegra2
dationofPlantCellWallPolysaccharides[J]1MicrobiologyandMolecularbi2ologyreviews,2001,65:49725221
[2]LUONTERIE,PAULAK,TENKANENM,etal1ActivityofanAsper2
gillusterreusa2arabinofuranosida1fionPhenolic2substitutedOligosaccharides[J]1JournalofBiotechnology,1999,67:412481
[3]OUSY,KWOKKC1ReviewFerulicAcid:PharmaceuticalFunctions,Prep2
arationandApplicationsinFoods[J]1JournaloftheScienceofFoodandAg2riculture,2004,84:1261212691
[4]KROONPA,GARCIA2CONESAMT,FILINGHAMIJ,etal1Release
ofFemlicAcidDehydrodimersfromPlantCellWallsbyFeruloylEsterases[J]1JSci1FoodAgile1,1999,79:42824341
[5]CREPINVF,FAULDSCB,CONNERTONIF1FunctionalClassification
oftheMicrobialFeruloylEsterases[J]1AppliedMicrobiologynology,2004,63:64726521
[6]MACKENZIECR,BILOUSD,,allolyticandxylanolyticenzyme]1AppliedandEnvironmentalMicrobiology,,53228391
[7]MATHEWS,ABRAHAMTE1ontheproductionofferuloylester2
asefromcerealbransandsugarcanebagassebymicrobialfermentation[J]1EnzymeandMicrobialTechnology,2005,36:56525701
[8]JOHNSONKG,SILVAMC,MACKENZIECR,etal1Microbialdegra2
dationofhemicellulosicmaterials[J]1Appl1Biochem1Biotechnol1,1989,20:245?2581
[9]TOPAKASE,VAFIADIC,CHRISTAKOPOULOSP1Microbialproduc2
tion,characterizationandapplicationsofferuloylesterases[J]1ProcessBio2chemistry,2007,42:497~5091
[10]DONAGHYJA,MCKAYAM1Productionofferuloyl/p2coumaroylesterase
activitybyPenicilliumexpansum,PenicilliumbrevicompactumandAspergillusniger[J]1JournalofAppliedBacteriology,1995,79:65726621
[11]ASTHERM,HAONM,ROUSSOSS,etal1FeruloylesterasefromAs2
pergillusnigeracomparisonoftheproductioninsolidstateandsubmergedfermentation[J]1ProcessBiochemistry,2002,38:68526911
[12]PANAGIOTOUG,GRANOUILLETP,OLSSONL1Productionandpar2
tialcharacterizationofarabinoxylan2degradingenzymesbyPenicilliumbrasil2ianumundersolid2statefermentation[J]1AppliedMicrobiologyandBiotech2nology,2006,27:1117211241
[13]DONAGHYJ,MCKAYAM1Purificationandcharacterizationofaferu2
loylesterasefromthefungusPenicilliumexpansum[J]1JournalofAppliedMicrobiology,1997,83:71827261
[14]CASTANARESA,MCCRAESI,WOODTM1Purificationandproperties
ofaferuloyl/p2coumaroylesterasefromthefungusPenicilliumpinophilum[J]1EnzymeandMicrobialTechnology,1992,14:87528841
[15]TOPAKASE,KALOGERISE,KEKOSD,etal1Productionandpartial
characterizationofferuloylesterasebySporotrichumthermophileundersol2id2statefermentation[J]1ProcessBiochemistry,2003,38:1539215431[16]CREPINVF,FAULDSCB,CONNERTONIF1Anon2modulartypeB
feruloylesterasefromNeurosporacrassaexhibitsconcentration2dependentsubstrateinhibition[J]1BiochemicalJournal,2003,370:41724271[17]GARCIA2CONESAMT,CREPINVF,GOLDSONAJ,etal1Theferu2
loylesterasesystemofTalaromycesstipitatus:productionofthreediscreteferuloylesterases,includinganovelenzyme,TsFaeC,withabroadsub2stratespecificity[J]1JournalofBiotechnology,2004,108:22722411[18]JOHNSONKG,HARRISONBA,SCHNEIDERH,etal1Xylan2hydrol2
ysingenzymesfromStreptomycesspp[J]1EnzymeandMicrobialTechnolo2gy,1988,10:40324091
[19]MUKHERJEEG,SINGHRK,MITRAA,etal1Ferulicacidesterase
productionbyStreptomycessp[J]1BioresourceTechnology,2007,98:21122131
[20]MATHEWS,ABRAHAMTE1Studiesontheproductionofferuloyles2
terasefromcerealbransandsugarcanebagassebymicrobialfermentation[J]1EnzymeandMicrobialTechnology,2005,36:56525701
[21]BONNINAE,BRUNELM,GOUYY,etal1AspergillusnigerI21472and
PycnoporuscinnabarinusMUCL39533,selectedforthebiotransformationofferulicacidtovanillin,arealsoabletoproducecellwallpolysaccharide2de2gradingenzymesandferuloyl]1EnzymeandMicrobialTechnolo2gy,2001,28:702801
[22,C,SANCHOAI,etal1Growth
ofBrewer’sspentgrainbyStrepto23339[J]1EnzymeandMicrobialTechnology,21441
]RUMBOLDK,BIELYP,MASTIHUBOVAM,etal1Purificationand
propertiesofaferuloylesteraseinvolvedinlignocellulosedegradationbyAu2reobasidiumpullulans[J]1AppliedandEnvironmentalMicrobiology,2003,69:5622256261
[24]BORNEMANWS,HARTLEYRD,MORRISONWH,etal1Feruloyl
andp2coumaroylesterasefromanaerobicfungiinrelationtoplantcellwalldegradation[J]1AppliedMicrobiologyandBiotechnology,1990,33:34523511
[25]BORNEMANWS,LJUNGDAHLLG,HARTLEYRD,etal1Purifica2
tionandpartialcharacterisationoftwoferuloylesterasesfromtheanaerobicfungusNeocallimastixstrainMC22[J]1AppliedandEnvironmentalMicrobi2ology,1992,57:3762237661
[26]MCDERMIDKP,MACKENZIERC,FORSBERGCW1Esteraseactivi2
tiesofFibrobactersuccinogenessubsp1succinogenesS85[J]1AppliedandEnvironmentalMicrobiology,1990,56:12721321
61阅读提醒您本文地址:
[27]FAULDSCB,WILLIAMSONG1Thepurificationandcharacterisationof42
hydroxy232methoxycinnamic(ferulic)acidesterasefromStreptomycesolivo2chromogenes[J]1JournalofGeneralMicrobiology,1991,137:2339223451[28]BAYEREA,CHANZYH,LAMEDR1Cellulose,cellulasesandcellulo2
somes[J]1CurrentOpinioninStructuralBiology,1996,8:54825571[29]TAMARUY,DOIPH1PectatelyaseA,anenzymaticsubunitofthe
Clostridiumcellulovoranscellulosome[J]1ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,2001,98:4125241291
[30]BLUMDL,KATAEVAIA,Li,etal1Feruloylesteraseactivityof
theClostridiumthermocellumcellulosomecanbeattributedtoprevi2ouslyunknowndomainsofXynYandXynZ1JournalofBacteriology12000,182:1346213511
[31]JOSEAM,TARBOURIECHN,SIMONJ,etal1TheStructureof
theFeruloylEsteraseModuleofXylanase10BfromClostridiumther2mocellumProvidesInsightsintoSubstrateRecognition[J]1Structure,2001,9:118321190

1
阿魏是什么 微生物阿魏酸酯酶的研究进展
?16 ?
2008No110
SerialNo1187
ChinaBrewing
SpecialDiscuss
我国名优黄酒的产生、特征、改革和发展
汪建国1,奕水明2
(11嘉兴酿造总公司,浙江嘉兴314000;嘉兴市质检所,浙江嘉兴314000)
摘 要:该文对黄酒主要香气、风味成分和工艺特征进行了分析、总结,并根据新时期消费者的口味结构变化,提出了对名优黄酒存在缺陷需进一步改革、创新和发展的思路。[www.61k.com)
关 键 词:名优黄酒;产生;特征;继承;发展
中图分类号:TS26214 文献标识码:A 文章编号:0254-5071(2008)10-0016-04
Emergence,characteristics,reformanddevelopmentofChinesefamoushigh2qualityricewine
WANGJianguo1,YIShuiming2
(11jiaxingBrewingGo1,Ltid1,Jiaxing31400,China;21JiaxingQualitySupervisionandInspectionInstitute,Jiaxing314000,China)Abstract:Inthispaper,theprize2winningbrandsofricewineaswellastheirquantityofawardsinallpreviousNationalConferencesofAlcoholicDrinkEvaluationwerebrieflyreviewed1Inaddition,themainaromaticcompounds,flavorcomponentsandprocessingfeaturesof2famousproductsand13highqualityproductsofricewinewereanalyzedandsummarized1Inthelightofthechangesofconsumertasteinthenewperiod,someapproachesforremovingtheexistingdefectsofricewinewereputforward1
Keywords:famoushigh2qualityricewine;emergence;characteristics;succession;development
我国黄酒的酿造历史悠久,其工艺成熟、技艺精良、品种繁多、风味独特,成为世界酒类上独树一帜的美酒。科学技术的进步、微生物学、酶化学的提高,总结各种传统生产工艺基础上,和创新的精神,质量,,极大地提高了产品质量和生产效率,推动了黄酒工业的快速发展,本文对黄酒的产生、特征、继承和发展作一简叙。1我国名优黄酒的产生与发展
黄酒的质量鉴别,主要是通过物理、化学分析和感官品质的方法评判。物理化学分析了解组成酒体的化学成分和物理状态。如酒精、总酸、糖分、总酯、氨基酸态氮等的含量以及甲醇、黄曲霉毒素B1、重金属等。但感官指标主要是通过人的感觉器官,依其多种黄酒的类型、品质、风格和它们之间的特征和风味差别来综合品评鉴别。
我国自1952年第一届评酒会开始到至今进行了五届,但对我国黄酒评酒只延续四届。每届评酒会不仅推动了生产发展,也起到了指导消费的作用。同时显示了不同历史
收稿日期:2007203212
2种国家名酒和13种国家优质黄酒以及1984年轻工业部颁布黄酒金杯奖8尊、银杯奖18尊和铜杯奖10尊。因此名优黄酒的生产遍及全国各省、市和地区,黄酒的品质和产量都有进一步提高,评酒标准和规定日臻完善。
第三届全国评酒会(1979年)开展了对绍兴黄酒总结、研究和提高,初步摸索到各类名优黄酒的生产规律和特征。特别是科学地总结了名优黄酒传统经验,去粗取精,肯定了传统工艺中的科学部分,改进了不合理的工艺,使名酒的生产发生了根本的变化,从师徒代代相传进入科学地指导名优酒生产。此外黄酒行业技术协作会广泛地开展了技术协作、交流经验、取长补短,使得名优黄酒的风格更加丰富多彩。并结合名优黄酒酿造用粮、糖化发酵用曲,工艺特征和类型进行充分酝酿,统一了主要品种、类型、风格典型的描述。111传统麦曲稻米黄酒(以绍兴黄酒为例)
色泽:橙黄色、清亮透明。香气:具有麦曲黄酒特有的香气,醇香浓郁。口味:(干型)具有元红酒特有口味,醇和、
作者简介:汪建国(19562),男,浙江嘉兴人,高级酿酒师,主要从事黄酒和调味品的研究工作。
[32]刘鹏展,欧仕益,包惠燕1阿魏酸酯酶的研究与应用[J]1食品研究与开
uloylatedoligosaccharidesfromsugar2beetpulpandwheatbranbyferulicacidesterasefromAspergillusniger[J]1CarbohydrateResearch,1994,263:25722691
[36]纵 伟,赵光远,张文叶1阿魏酸研究进展[J]1中国食品添加剂,2006,
(3):712741
[37]张 ,欧仕益,张 宁,等1酶解麦麸制备阿魏酸和低聚糖的研究[J]1
发,2006,27(5):16921721
[33]FAULDSCB,WILLIAMSONG1Releaseofferulicacidfromwheatbran
byaferulicacidesterase(FAEIII)fromAspergillusniger[J]1AppliedMi2crobiologyandBiotechnology,1995,43:1082210871
[34]DONAGHYJ,KELLYPF,MCKAYAM1Detectionofferulicacides2
teraseproductionbyBacillusspp1andlactobacilli[J]1AppliedMicrobiologyandBiotechnology,1998,50:25722601
[35]RALETMC,FAULDSCB,WILLIAMSONG,etal1Degradationoffer2
食品科学,2003,24(11):632681
[38]薛 枫,欧仕益,汪 勇,等1双酶法降解玉米麸皮制备低聚糖和阿魏酸
的研究[J]1粮食与饲料工业,2006(11):17219,26
61阅读提醒您本文地址:

1
61阅读提醒您本文地址:
二 : 生物技术进展论文
生物技术在鱼养殖中的应用
班级:A11行政1班 学号:110901108 姓名:张洪迪
摘要:随着科学技术的发展,除了传统的生物技术之外,分子技术的应用已经为提高世界水产养殖品种产量开辟了另一条途径。(www.61k.com)基因组信息库的增长,极大地提高了科技工作者对确认一些表现型基因的能力。转基因技术和目前尖端的技术:诸如DNA芯片技术、高分辨力的二维凝胶技术和高灵敏度的质谱仪等技术的发展联系起来,向水产养殖工作者展现出了一片美好的前景:包括生长率和经济效益的增加、抗病能力和抗冻能力的提高、养殖品种种质的改善以及通过改变它们的基因组成而创造出新的不同的品种。讨论了与功能基因组领域相关的技术以及它们在水产养殖领域中的应用。
关键词: 生物技术 鱼类养殖 潜在影响
我国水产养殖有着悠久的历史,远在3000多年前的殷末周初就有养鱼的记录,至公元前5世纪的春秋战国时代,陶朱公范蠡根据当时的养育经验编写了世界上第一部养育著作《养鱼经》。经过几千年的养鱼实践,不断积累了丰富的技术经验,水产养殖业取得了长足发展。到了近代,生物技术的全面发展,对水产养殖业产生了深远的影响,几乎对水产养殖的每个方面都起到了积极的指导作用。本文主要从鱼类遗传育种、养殖病害的检测和防治等方面简要论述生物技术的应用。
1 现代生物技术在鱼类养殖遗传育种方面的应用
1.1 多倍体育种技术 多倍体育种和雌核发育技术是在染色体水平上操作,进行品种的改良。目前,已取得良好的生产效益并显示出较好的产业化前景。 多倍体育种主要应用在促生长、提高品质、提高杂种成活率、保持杂种优势等方面。 通过温差处理、药物处理、水静压、杂交等诱导方法可产生多倍体。鱼类染色体的特点之一就是可塑性较大,易于加倍,如果在精子入卵而第二集体尚未排出卵外之时,对受精卵进行热休克(高温)、冷休克(低温)、高压或化学药品处理,组织第二极体排出卵外,则可诱导出三倍体个体。如果抑制第一次有丝分裂,则可获得四倍体个体。四倍体鱼再与二倍体染交,也可获得三倍体鱼。种间杂交受精卵经三倍体诱导技术处理而培养出的三倍体称之为杂交三倍体。三倍体鱼往往是不育的,这对经济鱼类的养殖意义重大,它避免了性腺发育阶段和产卵季节肉质下降及性腺发育时期的生长停止和死亡率上升,减少了养殖成本,利用其不育性还可以控制养殖鱼类的迅速繁殖和防止对天然资源的干扰。人工诱导的主要目的在于利用三倍体具有生长快、肉质好、生命周期长等特点,另外三倍体种群还具有较高的抗病力和抗逆性。
1.2雌核发育技术 雌核发育技术也是实现增产的重要手段。可用物理或化学因子处理授精前的精液,用带血的玻璃针激活卵子发育、杂交等人工诱导雌核发育,再经温差处理、秋水仙素和水静压等方法使之两倍化。雌核发育具有可建立纯系,可控制性别,且具有简便易行、快速可靠的优点。在海水动物中,由于性别不同而表现出生长速度不同是比较常见的生物学现象。目前,已经成功培育出红鲤(Cyprinus carpio)、斑马鱼(Hrachydanio recio)、尼罗罗非鱼(Oreachromis niloticus l)、莫桑比克罗鱼(oreachromis mossamb icus)等多个人工雌核发育品系,为研究鱼类性别决定机制、单性养殖及养殖新品种的开发研究和生产时间提供了几位宝贵的素材。
1.3转基因技术 在基因工程方面,转基因技术为水产生物遗传育种开辟了一条新途径。该项技术可以根据人们的需要,把有用的外源目的基因,如生长激素基因、抗冻基因、抗病基因等及其调控序列一起,导人受体的胚胎或受精卵中并使之表达,从而达到遗传改良
生物技术论文 生物技术进展论文
的目的。[www.61k.com)转基因鱼的研究工作在中国、美国、加拿大、日本、法国等已经展开,并取得了很大的成就。转基因技术的应用,为克服动植物种间杂交不育以及远缘杂交困难的问题,显示出极大的优势。目前在水产中的研究工作主要集中在鱼类。“全鱼”元件组成的转生长激素基因鱼散构建已在鲤鱼获得成功。养殖结果表明,转基因鲤鱼在生长速度和抗病能力上已显示出优势。从理论上说,利用转基因可以改变水生生物的遗传物质,使改进的优良性状得以遗传,进而从根本上长久的提高水产养殖的产量和改善产品的品质,所以其在水产养殖中的应用将随时间的推移而更加深入的推进水产养殖育种工作的研究。
1.4分子标记技术 分子遗传标记是指利用分子生物学方法来区分不同的个体或群体能够稳定遗传的物质或性状,是生物个体或群体间遗传差异的客观表征。目前限制性片段长度多态性(RFLP),简单序列长度多样性(sSLP),随机扩增多态性DNA(RAPD),单核苷酸的多态性(SNP)是最流行的分子标记技术。分子标记一个重要的应用就是来构建遗传图谱。遗传图谱的建立为基因定位,特别是一些重要经济性状和数量性状位点的定位,并为最终克隆这些基因提供了基础,同时还可以推动生物的分子标记辅助选育和遗传改良。分子辅助育种已成为有效的育种工具,它是根据与某一性状或基因紧密连锁的标记的出现来推断该基因或性状的有无,从而进行选育的方法,可以增加选择的准确性,大大缩短育种的周期。进而大幅度提高中国养殖生产中优质原良种的普及率,达到增产增效的目的。Gomez-Raya 等认为在进行的选择育种工作中如果采用了分子辅助育种技术,那么育种速度将提高大约11%。例如梁利群等(2000)利用RAPD技术对荷包红鲤(Cyprinus carpio Linnnaeus)抗寒品系的基因组DNA进行分析,找到了用RAPD技术从分子生物学角度对荷包红鲤抗寒品系进行鉴定的方法。
2现代生物技术在鱼类养殖病害的检测方面的应用
2.1核酸探针技术 核酸探针技术通过分析病原生物的遗传结构,人们发现各种病原体均有其独特的基因结构。根据这一段基因结构的序列,就可以设计可与其互补的DNA 探针,与病原生物的DNA进行杂交,以此来确定病原携带者和传播者的一种分子生物学技术。该技术具有灵敏度高、特异性强等优点,近年来在对虾病毒病的检测中倍受青睐。美国科学家针对IHH-NV(传染性皮下和造血组织坏死病毒)研制的点杂交核酸探针已经商业化生产。中国科研人员针对对虾造血组织坏死杆状病毒研制的核酸探针,并将之用于检测受造血组织坏死杆状病毒感染的对虾,取得了满意的结果。
2 .2PCR技术 PCR技术即聚合酶链式反应,又称为 DNA体外扩增技术,其原理是在了解了病原生物特殊的遗传结构的基础上,设计对专性引物再通过PCR扩增产生出电泳可见的扩增产物,从而通过电泳、染色把它显示出来。该方法既具有DNA探针方法的灵敏度,又避免了昂贵且危险的同位素的使用,并可以更快得到诊断结果,在对虾病原检测方面,中国研究人员已将这一技术成功地用于对虾病毒病和细菌病的检测。目前,很多国家均已开发出商品化的造血组织坏死杆状病毒的PCR检测试剂盒。另外,PCR技术与其他技术相结合开发出新的PCR技术更显示出其独特的优越性,如荧光定量PCR技术、PCR-ELISA技术等。现均已应用于水生动物疾病的诊断中,并已显示出巨大的潜力及广阔的应用前景。
2.3单克隆抗体技术 单克隆抗体技术是近年出现的重要技术之一。它是在Milstein发明的杂交瘤技术基础上建立的。单克隆抗体与体外侵物的结合具有很高的敏感性和检测性,因而可作为诊断技术。单克隆抗体技术在对养殖对虾暴发性流行病的病原及传播途径的调查中,发挥了重要作用。应用这一技术,能对养殖对虾的发病情况提前20--40天作出预报,为生产上及时采取对应措施提供了可靠的依据。另外,单克隆抗体技术可以与其它免疫技术相结合,如放射免疫分析技术、酶联免疫检测技术、间接荧光抗体技术等,进而对疾病的检测做到更加准确、迅速。
生物技术论文 生物技术进展论文
2.4基因芯片 基因芯片又称DNA芯片(DNA chip)或DNA微阵列(DNAmieroarray),是将大量基因探针,按特定的排列方式固定在固体载体上,它通过与被检测基因的碱基序列进行固定互补匹配,实现核酸序列的分子识别,是高效的大规模获取相关生物信息的重要手段。(www.61k.com)与传统核酸印迹杂交技术相比,基因芯片具有高效、快速和大规模平行处理的优点,是在传统生物技术和疾病诊断等方面的创新和飞跃。基因芯片在DNA测序和基因表达分析、临床诊断、药物和疫苗研究等领域有极大应用前景。针对病原微生物基因组的特征性片段、染色体DNA的多态性、基因变异的位点及特征等,设计和选择合适的核酸探针并制成芯片,通过与样品中提取的核酸(DNA或RNA)杂交,一步检测就能获得病原微生物种属、亚型、毒力、抗药、致病、同源性、多态型、变异、表达以及感染病原微生物相对数量等相关信息,为疾病的诊断和治疗提供了一个很好的切入点,也为平行研究病原微生物的大量基因组和功能组提供了有利的工具。
3现代生物技术在鱼类养殖病害的防治方面的应用
3.1基因工程疫苗 基因工程疫苗的开发基因工程疫苗是将病毒中负责转录抗原蛋白质的基因片段分离出来,与载体结合成重组DNA,然后转人细菌、酵母菌、病毒或动物细胞的DNA中。在这些寄生生物的增殖过程中,借基因的表达,大量生产抗原蛋白质作为疫苗使用。疫苗的功用在于能诱导动物对某些特异的致病菌产生抗体,增强抵御病原侵袭的能力。应用基因工程技术研制疫苗,其优点在于抗原专一,避免了常规疫苗将整个病原基因组转入接种鱼而造成可能的病原体传播和危害。其次,基因工程疫苗可通过基因剪接将不同病原体的致病性抗原决定簇的编码基因拼接在一起而构建表达复合疫苗。第三,这种转基因疫苗可用发酵技术进行工厂化生产,易于生产出价格低廉、稳定性好的产品。最近,科学家否定了无脊椎动物没有免疫特异性的说法,指出无脊椎动物不仅具有免疫特异性,而且对抗原具有记忆能力,这预示着基因工程疫苗的开发和使用将在水产养殖中病害的防治中起到更加重要的作用。
3.2免疫刺激剂 免疫刺激剂的开发免疫刺激的研究和开发是近年来蓬勃兴起的一个新的研究领域。免疫刺激剂能诱导和刺激水产养殖动物自身的非特异性免疫系统与各种病原作斗争,通过增强自身的免疫能力,达到预防和减轻疾病的目的。在甲壳动物中当外界病原侵入时,甲壳动物可产生外源凝集素,这种物质与病原微生物细胞壁上的糖类结合,使其更易于被白细胞消化和杀死。已研制出来的几种非特异性免疫刺激剂在白对虾和斑节对虾的抗病试验中都取得了理想的效果。
3.3荧光抗体技术 荧光抗体技术是根据抗原具有高度特异性的反应,吧荧光素作为矿院标记物,在荧光显微镜下减产呈现荧光的特异性抗原复合物及其存在部位,在水产动物病原体的检测上得到了一定的应用。鄢庆枇应用荧光抗体技术检验牙鲆体内的弧菌。该技术的主要优点是特异性强、速度快、灵敏度高,但也存在缺点如非特异染色问题难以完全解决,造作程序较繁琐,需要特殊的昂贵仪器和染色体本不能长期保存等。
4现代生物技术在鱼类养殖营养与饲料方面的应用
4.1基因工程技术 随着基因工程技术的发展和运用,很多有生物活性的多肽或蛋白质可以较容易地被合成,且成本低廉。由于鱼类胃肠道对多肽和蛋白质的吸收能力较强,使得基因重组产品能较方便、快捷地对鱼类的生理活动进行调控,尤其是在促进鱼体生长方面。例如:对银大麻哈鱼、草鱼投以基因重组生长激素,可有效刺激鱼体成长。
4.2酶工程技术 酶工程是生物技术的一个重要组成部分,它是指在一定的生物反应器内,利用酶的催化作用,进行物质转化的技术。而酶制剂则是酶3工程的代表产物。酶制剂室从动、植物和微生物中提取制备的具有酶专一的高效生物活性物质,通常与少量载体混
生物技术论文 生物技术进展论文
合而制成粉剂。[www.61k.com]目前,应用生物技术生产的酶有蛋白酶、脂肪酶、糖化酶、植酸酶、纤维素酶等。大多数酶来自于真菌类。水产动物饲料中添加适量合适的酶制剂,有利于促进养殖动物对饲料的吸收、转化,提高养殖动物的生产性能和健康水平。
4.3微生物工程技术 微生物工程也称发酵工程,是一门将微生物学、生物化学和化学工程学的基本原理有机地结合起来,利用微生物的生长和代谢活动来生产各种有用物质的工程技术。该技术可用于饲料的育藏、改善饲料的营养价值、微生物饲料的生产以及用微生物来净化有毒的物质,消除有毒气体和恶臭物质以及处理有机废水、废渣等;还可用于生产抗生素、维生素、透明质算、微生物蛋白、氨基酸和一些食品添加剂(如柠檬酸、乳酸、天然色素等)等。饲料中添加氨基酸可以平衡氨基酸的比例,提高蛋白质的利用效率,减少氮排出造成的环境污染。
现代生物技术是20世纪末人类科技史中最令人瞩目的高新技术之一,同时也是21世纪世界各国科学技术发展的重要内容。现代生物技术的应用为鱼类养殖业的发展带来了革命性的变化,从鱼类遗传多样性、种质资源、新品种培育、免疫预防、疾病诊断、水质监测等各个方面无一不渗透现代生物技术的应用.。然而其在生产上发挥更大的作用,还有待于现代生物技术更深一步的发展。相信随着试验方法的不断优化和设备的日益更新,生物技术的发展将使鱼养殖走上可持续发展的道路。但在我国生物技术还是一门年轻的学科,需要加强研究力度。另外,目前我国的鱼养殖业基本上还处于传统型,要实现水产养殖业的现代化,有赖于海洋生物技术的发展和应用。可以预言,对世纪是海洋的世纪,在合理开发和利用海洋生物资源,发展渔业经济和保护渔业环境中,海洋生物技术是大有可为的。
参考文献
1.刘晓慧,王义权,分子标记技术在水产动物研究中的应用。厦门大学学报。2008,47(2):228-231
2.梁利群,孙效文。用RAPD技术鉴定荷包红鲤抗寒品系。水产学杂志,2000,13(2):15-20
3.李卢,人工雌核发育在鱼类遗产育种中的应用。水产科技情报,2006,33(2):56-58
4.邹为民,陈信廉.细菌自动鉴定仪在水产动物疾病诊 断中的应用研究 [J] .珠江水产,2000, (4):9~11.
5.金丽.鱼类的人工繁殖技术 [J] .生物学教学,2005,30 (4):2~4
6.赵浩斌,陈尚萍,朱作言.转基因鱼的研究进展 [J] .农业生物技术学报,1999 (3):30l~306
7.温茹淑,崔淼.转基因鱼的研究进展 [J] .嘉应大学学 报,2002 (6):40~45
8.陈光荣,肖克宇,翁波,等.鱼类细胞工程遗传育种 研究进展,水利渔业,2004,24 (1):4~6
9.严绍颐.鱼类细胞核转植 (基因组转移) 和育种 [A] . 海洋生物技术研讨会论文集 [C],1991,79~83
10.谢忠明.“八五”国家重点推广养殖对象:颖鲤 [J] . 淡水渔业,1992, (2):48
三 : 乌鳢(♂)×斑鳢(♀)杂交繁育及杂交子一代生物学研究进展

四 : 30GIS技术在物流管理中的应用及研究进展
第30卷 第6期2008年12月
武汉理工大学学报?信息与管理工程版
JOURNALOFWUT(INFORMATION&MANAGEMENTENGINEERING)
Vol.30No.6
Dec.2008
文章编号:1007-144X(2008)06-0982-05
GIS技术在物流管理中的应用及研究进展
郭建宏,欧阳钟辉
(泉州师范学院陈守仁工商信息学院,福建泉州362000)
摘 要:对地理信息系统(GIS)技术在物流管理中的应用及研究进行了综述,包括基于GIS的物流中心选址、物流配送车辆定位导航和物流配送系统集成。其中说明了GIS在物流配送的实时调度、位置选择等方面有很好的应用前景,但受系统开发成本高、专业性强和通用性差等因素作用,。指出了标准化、集成化和网络化是GIS物流行业应用的发展方向。关键词:地理信息系统;物流管理;选址;定位导航;中图法分类号:TP368.6 文献标志码:A
、算机系统。一个与数字化经济相适应的现代化物流配送系统涉及到多方面的因素,如配送中心位置的选择、配送点的布局、配送路径优化、配送车辆的实时监控与调度和配送服务质量的提升等,这些因素都与地理位置密切相关。物流信息具有空间尺度、空间特征的性质,物流信息中大约80%的信息与空间位置有关,而运输、仓储和配送等涉及到的信息几乎全部与空间位置有着直接的
[124]
关系,决定了它与以GIS为核心内容的空间信息技术的集成成为可能。
性和定量两大类。定性的方法主要采用层次分析法和模糊综合评价相结合对各个方案进行指标评价,找出最优地址。定量的方法主要包括重心法、运输规划法、Cluster法、FLP法、Baumol-Wolfe法、混合0-1整数规划法、双层规划法和遗传算
[7-11]
法等。总体来看,上述各种模型都存在以下问题:①模型常常假设需求量集中于某一点,而实际上需求来自分散于广阔区域内的多个消费点。②运输费用通常假设运价随运输距离成比例增加,然而,大多数运价是由不随运输距离变化的固定部分和随运输距离变化的可变部分组成的。③模型中物流中心与其他网络节点之间的路线通常假定为直线,实际上这样的情况很少。
利用GIS能较好地解决上述问题,在GIS中供应地和需求地都是空间实体,根据其实体特征可用点来表示,同时整个计划区域的基本信息可通过空间数据表达,分别对应着空间特征数据、属性特征数据和拓扑特征数据。VLACHOPOULOU等结合GIS网络分析的特点,提出了一种基于
[13]
GIS的配送中心最优设置分析模型。陈曦等对GIS环境下的物流配送中心选址模型与算法进行了研究,设计了遗传算法与线性规划相结合
[14]
的计算方法。贺政纲等论述了利用GIS选址的关键技术,提出了基于GIS的最短路径算法的
[15]
实现方法。宣登殿等利用GIS
的空间分析能
[12]
1 基于GIS物流中心的选址研究
物流中心选址是物流系统中具有战略意义的投资决策问题,对整个系统的物流合理化和商品流通的社会效益有着决定性的影响。但由于商品资源分布、需求状况、运输条件和自然条件等因素的影响,使得即使在同一区域内的不同地方建立物流中心,整个物流系统和全社会经济效益也是不同的。因此,物流中心的选址应以物流系统和社会的经济效益为目标,用系统学理论和系统工程的方法,综合考虑多种因素,对物流中心的位置进行科学的研究和决策。
物流配送中心选址研究的方法大致可分为定
[526]
收稿日期:2008-06-05.
作者简介:郭建宏(1973-),男,甘肃定西人,泉州师范学院陈守仁工商信息学院博士.
基金项目:福建省青年科技人才创新基金资助项目(2005J056);泉州市科技计划资助项目(2006G8).
第30卷 第6期郭建宏,等:GIS
技术在物流管理中的应用及研究进展
983
力,采用克理斯塔勒六边形法则划分区域的方法,有效解决了配送中心数目的确定问题,并建立了
[16]
配送中心空间布局优化模型。张席洲等讨论了GIS的空间查询、空间分析和网络分析等功能在物流中心选址中的应用,总结了利用GIS选址的实现步骤。
基于GIS的选址能摆脱数学模型的一些不足之处,充分考虑了实际客观情况,通过对空间信息和非空间信息同时进行分析和处理,使之得到的物流中心的位置更加准确,能更符合实际要求,因而它具有很高的实际应用价值。
量变化情况,制定最佳行车计划等。
3 基于GIS的物流信息系统的集成
物流是一个大范围的活动,物流信息贯穿于物流活动的全过程。物流信息具有信息源点多、分布广、信息量大、动态性强、信息的价值衰减速
[27]
度快和及时性要求高等特性。这意味着物流信息的收集、加工和处理要求速度快、难度大。目前的物流管理系统主要还是在MIS的基础上将数据用报表、图表等形式表达出来,尽管相对于原来单纯的文字形式更易理解,但信息仍然过于繁锁,信息的理解、,物流,无法。随着物流、智能化和集成化方向,物流信息系统对空间信息的表达以及物流
[28]
可视化的需求愈来愈强。
在应用GIS技术进行物流信息管理时,需要对异构数据进行处理。这就要通过GIS技术将不同类型的信息系统进行集成,从异构的系统获取有关信息,对信息资源进行合理共享和优化利用。国内外有许多学者把研究集中在GIS系统集成模
[29-32][33][34]
式和集成方法上。郑长江、卢战伟等对GIS与物流信息系统数据集成和功能集成技术进行了研究,认为目前在数据集成方面比较理想的方式是,在同一RDBMS中集成空间数据、非空间数据和影像数据。在功能集成方面认为COR2BA技术是当前解决分布、异构问题的有效技术,而Agent具有自主性、交互性、反应性和主动性等特点,采用这种技术对于实现分布式信息系统在网络通信和协作处理方面有很大优势,可较好地解决系统数据共享和服务共享,便于子系统间的应用协作和智能化服务,并指出Agent技术目前还处于理论研究阶段。
也有学者侧重于基于GIS物流系统模型构造
[35]
的研究。胡志杰等研究了基于ArcGIS的物流
[36]
配送系统集成的总体框架。楼月明等提出了基于WebGIS的物流配送系统模型,并利用J2EE多层体系,采用结构灵活的4层结构构建了基于WebGIS配送系统模型的体系结构。刘志强等
[37]
2 GIS在物流车辆定位导航中的应用
基于GIS的物流车辆定位导航一直是国内外学者研究的热点问题,也是以GIS。2
[17]
BERG[18]
DEREKENARIS等集成了基于GIS、GPS(全球定位系统)和GSM(全球移动通信系统)(即3G技术)的急救车辆导航管理系统。屈晓翔等对基于GIS开发的物流配送车辆监控导航系统进行了需求分析,包括综合查询、配送订单排序及静态线路优化、动态车辆监控导航等,提出了系统功能模块设计方案,包括线路优化模块、综合查询模块、与车辆终端的交互模块、车辆监控及导航模块和地图操作模块等。张
[20]
贵明介绍了GIS在车辆导航系统中最佳路径算法,分析了算法的详细步骤和算法复杂度。夏
[21]
红霞等介绍了一种基于GIS和GPS集成的移动车辆综合服务系统,设计了其监控中心模块的部分功能,讨论了在监控中心模块设计中的部分
[22]
关键技术。张剑等提出一个基于GPRS通信平台,利用WebGIS实现车辆定位导航信息分布式发布的功能框架,并就GIS信息在GPRS移动终端上的无线发布作了进一步探讨,提出了相应解决方案。
目前物流车辆的定位导航大多是基于GIS电子地图查询的,其智能化、自动化程度还不高。现代物流企业要注重自身GIS、GPS建设的同时,还应与交通、公安等部门的相关系统进行衔接,充分利用他们的资源,达到事半功倍的效果,目前这两个部门的GIS、GPS建设走在了物流企业的前[23-26]面。如交通部门的智能运输系统(ITS)与GIS-T(GISfortransportation)可为物流管理提供数字化平台,可选择、优化运输路线,分析货物流
[19]
提出了基于3G技术的物流配送信息系统的模
型,开发了以订单为中心的物流库存控制、配送控制和车辆在途定位监控的系统模块。陈佳娟[38]
等采用J2EE技术和GIS技术设计了面向第三方物流企业的B2B和B2C实时配送管理系统,分
984
武汉理工大学学报?信息与管理工程版2008年12
月
析了系统功能结构、网络拓扑结构、系统配置方法
以及配送路网制作方法。
由于物流活动几乎涉及到商务活动的各个行业,有不少研究是针对GIS在物流各行业的信息系
[39]
统的研究。SONG等设计了基于GIS的报纸配
[40]
送系统;BUTLER等介绍了将空间信息技术集成到牛奶收配系统的数据结构和实现方法;孙东[41]
辉论述了GIS技术在邮政物流信息系统中的应用;嵇芸芸等讨论了基于GIS城市废弃物物流
[43]
管理信息系统的开发;郭建宏等研究了城市水果物流分区配送理论和方法,实现了基于GIS的城市水果收配一体化决策支持系统,有效解决配送车辆实时调度和定位导航问题。另外,针对物流配送的各个环节,一些机构和单位也开发了很多基于GIS的物流系统,如物流配送空[44248]492统、2]
。
,国外物流企业已经形成了以系统技术为核心,以信息技术、运输技术、配送技术、装卸搬运技术、自动化仓储技术、库存控制技术和包装技术等专业技术为支撑的现代化物流管理格局。在美国仅配送相关软件就有24种之多,并且无一例外地选择了GIS作为实现路线和站点可视化的基础平台,物流管理系统从当初为满足某一目的的管理系统逐渐向综合物流信息平台过渡。我国在物流软硬件方面也开始了深入的研究,立体仓库、GPS定位技术逐步推广开来,不少机构和企业从不同的角度出发,针对不同的需求对象以及应用范围,构建了相应的基于GIS的物流配送系统或信息平台。我国最大的国际货运代理集团中外运,业务涉及空、海、汽和国际联运等领域,由于采用了基于WebGIS物流配送管理系统,因此,实现了利用GPS进行物流配送全程实时跟踪,客户通过因特网登录中外运网站,输入运单号就可以查出货物的位置、运输轨迹、当前位置和状态,方便客户随时监视自己的货物。我国4大海外投资集团之一的招商局集团所属的招商迪辰公司是国内规模最大的物流供应链服务企业,长期致力于推动物流与GIS技术发展和应用研究,开发的基于GIS/GPS的运输管理系统具有车辆实时精确定位、历史行程回放、物流资源查询和车辆信息管理等强大功能,已用于厦门邦辉等多家高科技物流企业,并为香港最大的一家物流公司装配了“路路通”系统。还有以“零库存”闻名的海尔集团,采用GIS技术实现JIT(justin
[54]
[42]
time)过站式物流配送模式,提出“在途”就是“库
存”的理念,实施了“以时间消灭空间”的业务流程再造。目前海尔物流已逐渐从企业物流向现代物流企业转变,不定期地承接美日等多家外资企业的物流总代理,同时开展了对外物流咨询、信息系统设计和物流知识培训等业务。另外,部分GPS提供商、凡丰物流、广东快步易捷公司等推出了一些GIS/GPS/GSM集成的物流软件。但是由于我国基于GIS的物流研究尚处于摸索阶段,因而还缺乏一个公认的优化解决方案。
4 GISGI,随着其结从整个未来软件应用模式的发展趋势看,基于GIS的物流软件设计和应用必然会向着标准化、集成化以及网络化等方向发展。
标准化就是要结合和借鉴国际与国内现有GIS标准化、物流标准化进程,充分研究GIS物流行业应用需求,制定物流地理信息系统(GIS-L)
基础性和方法性规范,同时促进物流标准化进程。可以说物流标准化和GIS-L标准化是GIS物流行业应用的基础。
基于GIS的物流软件开发和应用要充分利用现代先进的网络技术、空间数据库和组件技术等,构建具有标准接口的、适应性强的和随需而变的灵活的基础架构,使基于GIS的空间信息和非空间信息以及方法和模型能够无缝集成,使系统构建从低层次的软件开发向高层次的集成化过渡。
同时,在基于GIS的物流软件开发应用中,要充分利用现代网络技术,实现物流信息的数字化、网络化,完成物流信息资源充分开发和普遍共享,以降低物流成本,提高物流效率。另外,网络化不能局限于企业个体的网络化,要注重基于GIS的公共物流信息平台建设,推进GIS行业应用。公共物流信息平台可以利用其庞大的资料库以及开放性功能实现企业自身的信息交流、位置选择、定位导航和空间决策支持等的信息化管理。可以说,基于GIS公共物流信息平台是物流行业信息化的捷径。参考文献:
[1] 王 健.现代物流概论[M].北京:北京大学出版
第30卷 第6期
社,2005.
郭建宏,等:GIS
技术在物流管理中的应用及研究进展
985
[18] DEREKENARISG,GAROFALAKISJ,MAKRISC,etal.
IntegratingGIS,GPSandGSMtechnologiesfortheef2fectivemanagementofambulances[J].Comput.,En2viron.andUrbanSystems,2001,25(5):267-278.
[19] 屈晓翔,张汉江,赖明勇.GIS在物流配送车辆监控
[2] COYLEJ,BARDIE,LANGLEYJJ.企业物流管理
[M].7版.文 武,译.北京:电子工业出版社,2004.
[3] CHRISTOPHERM.物流与供应链管理[M].何明
珂,译.北京:电子工业出版社,2006.
[4] HESSEM,RODRIGUEJP.Thetransportgeographyof
logisticsandfreightdistribution[J].JournalofTrans2portGeography,2004(12):171-184.
[5] GOETSCHALCKXM,VIDALCJ,DOGANK.Model2
inganddesignofgloballogisticssystems:areviewofintegratedstrategicandtacticalmodelsanddesignal2gorithms[J].EuropeanJournalofOperationalRe2search,2002(143):1-18.
[6] 冯耕中.物流配送中心规划与设计[M].西安:西安
导航信息系统中的应用[J].系统工程,2005,
23(6):39-45.
[20] 张贵明.GPS/GIS车辆导航系统中最佳路径算法
研究[J].四川师范大学学报:自然科学版,2005,
28(4):497-500.
[21] 夏红霞,周 宏,杨红云,等.基于GIS/GPS车辆监
控系统实现及关键技术[J].微机发展,2004,14
(8):100-102.
[22 张,GIS在GPS车辆定
J,2004,
)-.
].GPS与GIS及其在现代物流中的应用
[J].物流技术,2002(4):12-14.
[24] THONGCM,WONGWG.UsingGIStodesigna
trafficinformationdatabaseforurbantransportplan2ning[J].Comput.,Environ.andUrbanSystems,1997,21(6):425-443.
[25] MILLERHJ.GISsoftwareformeasuringspace-time
accessibilityintransportationplanningandanalysis[J].GeoInformatica,2000,4(2):141-159.
[26] GOODCHILDMF.GISandtransportation:statusand
challenges[J].GeoInformatica,2000,4(2):127-139.
[27] 牛东来.现代物流信息系统[M].北京:清华大学
交通大学出版社,2004.
[7] 付鹏程,刘金华.[J].82.
[8] 杨 波,,.数学模型[J].,2002,10(5):57-61.
[9] 魏光兴.物流配送中心选址方法综述[J].物流与交
通,2005(9):48-49.
[10] ELONSC,MARIATAS,ALEXAF,etal.Age2
neticalgorithmforsolvingacapacitatedP-medianproblem[J].NumericAlgorithm,2003,35(4):373-388.
[11] LAPORTEG.Thevehicleroutingproblem:anover2
viewofexactandapproximatealgorithms[J].Europe2anJournalofOperationalResearch,1992(59):345-358.
[12] VLACHOPOULOUM,SILLEOSG,MANTHOUV.
Geographicinformationsystemsinwarehousesitese2lectingdecisions[J].InternationalJournalofProduc2tionEconomics,2001,71(1-3):205-209.
[13] 陈 曦,傅 明.GIS环境下的物流配送中心选址
出版社,2004.
[28] 刘志强,丁 鹏,盛焕烨.物流配送系统设计[M].
北京:清华大学出版社,2004.
[29] 闾国年,张书亮,龚敏霞,等.地理信息系统集成原
理与方法[M].北京:科学出版社,2003.
[30] 陈述彭,鲁学军,周成虎.地理信息系统导论[M].
模型与算法研究[J].计算技术与自动化,2001,
20(4):6-9.
[14] 贺政纲,帅 斌,廖 伟.GIS在物流中心选址中的
北京:科学出版社,2005.
[31] CSERL,CSELENYIJ,GEIGERM,etal.Logistics
fromIMStowardsvirtualfactory[J].JournalofMate2rialsProcessingTechnology,2000(103):6-13.
[32] CHOUINARDM.Integrationofreverselogisticsac2
tivitieswithinasupplychaininformationsystem[J].ComputersinIndustry,2005(56):105-124.
[33] 郑长江,王 炜,朱 成.GIS与物流信息系统集成
应用[J].交通标准化,2004(5):17-20.
[15] 宣登殿,胡大伟,蔺宏良.基于GIS的城市物流配
送系统规划方法[J].长安大学学报:自然科学版,
2006,26(2):84-87.
[16] 张席洲,龚奇才.基于GIS的物流中心选址[J].物
流技术,2005(10):249-252.
[17] HAFBERGG.
Integrationofgeographicinformation
systemsandnavigationsystemsformoving(dynamic)objectslikevehiclesandships[C].ProceedingsoftheESRIUserConference.[s.l.]:[s.n.],1995:1980-1988.
技术研究[J].交通与计算机,2005,23(2):
59-63.
[34] 卢战伟,霍 亮,钱曾波.GIS与城市物流配送集成
技术的研究[J].交通科技,2003(3):84-86.
[35] 胡志杰,王长琼.基于ArcGIS的物流配送系统集
成研究[J].物流技术,2005(3):35-37.
986
武汉理工大学学报?信息与管理工程版2008年12
月
[36] 楼月明,李 勃.基于WebGIS的物流配送系统的[45] GAYIALISSP,TATSIOPOULOSIP.DesignofanIT
-drivendecisionsupportsystemforvehicleroutingandscheduling[J].EuropeanJournalofOperationalResearch,2004(152):382-398.
[46] KARAGEORGOSA,MEHANDJIEVN,WEICHHART
G,etal.Agent-basedoptimizationoflogisticsandproductionplanning[J].EngineeringApplicationsofArtificialIntelligence,2003(16):335-348.
[47] 李 军,曹立明,王小平.基于GIS的物流配送路
研究[J].云南地理环境研究,2005,17(增刊):
61-62.
[37] 刘志强,周 枫.基于3G的物流配送系统设计与
实现[J].计算机工程与应用,2002(18):
249-251.
[38] 陈佳娟,王云鹏,纪寿文.第三方物流实时配送管
理系统的架构及路网GIS显示设计[J].计算机应用与软件,2004,21(10):30-32.
[39] SONGSH,LEEKS,KIMGS.Apracticalapproach
tosolvinganewspaperlogisticsproblemusingadigit2almap[J].Computers&IndustrialEngineering,2002(43):315-330.
[40] BUTLERM,HERLIHYP,KEENANPB.Integrating
informationtechnologyandoperationalresearchinthemanagementofmilkcollection[J].ofEngineering,2005(70):[41] 孙东辉.[J].5)17-19.
[42] 嵇芸芸,,曾 鹏.基于GIS的城市废弃物
径计算[J].计算机应用与软件,2004,21(10):
12-14.
[48] 戢一鸣.基于GIS的现代物流系统配送算法的研
究与应用[J].:信息与管理工2004,(.
[49,S的可视物流管理
].,2005(5):11-15.
],刘丙午,刘 军.信息可视化技术在物流
信息系统中的研究与应用[J].物流科技,2005,
23(8):22-25.
[51] 王成金.GIS技术在物流企业信息平台中的应用研
物流管理信息系统的开发与研究[J].物流技术,
2005(9):181-183.
[43] 郭建宏,钱莲文,欧阳钟辉,等.基于GIS的城市水
究[J].商业研究,2005(19):205-207.
[52] 石 晶,刘晓馨,段 敏.GIS/GPS技术在物流园
区信息平台中应用研究[J].交通企业管理,2006
(6):32-33.
[53] 霍 亮.基于Agent技术的空间物流信息系统框架
果物流分区配送辅助系统研究[J].中南林业科技大学学报,2007,27(4):119-122.
[44] TARANTILISCD,KIRANOUDISCT.Usingaspa2
tialdecisionsupportsystemforsolvingthevehicleroutingproblem[J].Information&Management,2002(39):359-375.
结构研究[J].测绘通报,2006(9):26-30.
[54] 中国物流与采购联合会.中国物流年鉴[M].北
京:中国物资出版社,2006.
ApplicationandResearchProgressofLogisticsManagementBasedonGIS
GUOJianhong,OUYANGZhonghui
Abstract:TheapplicationandresearchprogressoflogisticsmanagementbasedonGeographicInformationSystem(GIS),inclu2dingtheselectionoflogisticscenter,thelocationandnavigationofvehicleandtheintegrationofdistributionsystemwerereviewedaccordingtorecentreferences.Althoughtherearegoodapplicationsinthefieldofreal-timeschedulingdistributionvehicleandselectinglogisticscenter,yettheextensiveapplicationofsystemcannotworkbecauseofthelimitedhighdevelopmentcostandpoorgenerality.Itwaspointedoutthatstandardization,integrationandnetworkwillbethefuturetrendofGISforlogisticsapplica2tion.
Keywords:GeographicInformationSystem(GIS);logisticsmanagement;selectioncenter;locationandnavigation;systeminte2gration
GUOJianhong:Doctor;TSLSchoolofBusinessInformationTechnology,QuanzhouNormalUniversity,Quanzhou362000,China.
[编辑:王志全]
五 : 植物凝集素的研究进展
第48卷第2期
2003年4月
武汉大学学报(理学版)
J.W uhan Univ.(Nat.Sci.Ed.)
V0_-48 No 2
ADr.2002.232~ 238
文章编号:0253—9888(2002’)02—0232—07
植物凝集素的研究进展
梁 峰,常团结
(商丘师范学院生物系,河南商丘476000)
摘要:综述了有关植物凝集紊的研究进展,介绍了植物凝集紊的分类、糖结合特性、近年来有关植物凝集隶
蛋白晶体结构的研究,以及其与糖结合能力相关的生物学功能.
关键 词:植物凝集紊{糖结合活性;晶体结构;功能{抗虫植物基因工程 中图分类号:Q 946.L 文献标识码:A
植物凝集素是一类具有高度特异性糖结台活性
的蛋白,存在于许多植物的种子和营养器官中.第
一个植物凝集素是在蓖麻籽中发现的,到目前为止,
已经发现了多种不同的植物凝集素,在生理生化及
分子生物学方面对它们进行了许多研究,特别是近
年来,克隆了许多植物凝集素基因,对多种植物凝集
素进行了蛋白质晶体结构的研究,对植物凝集素的
生物学功能有了更深的认识.随着抗虫植物基因工
程的研究进展,植物凝集素在抗虫基因工程上的应
用受到了越来越多的重视,获得了许多转植物凝集
素基因的抗虫转基因植物.本文介绍植物凝集素近
年来的研究进展及其应用.
1 植物凝集素的发现
植物凝集素最早发现于1888年,Stillmark在
蓖麻(Ricinus COYT"t1"YtU~ti$L.)籽萃取物中发现了一
种细胞凝集因子,它具有凝集红细胞的作用.以后
多种植物凝集素不断得到发现,在单子叶和双子叶
植物中都发现有植物凝集素的分布,存在于许多植
物的储藏器官和繁殖器官中.植物凝集素长期被认
为是一种典型的种子蛋白,这是由于许多凝集素是
在种子里发现的.现在许多植物营养器官中均发现
了凝集素,非种子来源的凝集素和种子来源的凝集
素同样分布广泛.植物凝集素在种子、营养器官中
的丰度、分布的部位不尽相同.例如,种子来源的凝
集素主要分布在子叶、胚乳和胚轴中.非种子来源
的凝集素在营养器官如叶片、茎、茎皮、鳞茎、块茎、
球茎、根状茎、根、果实、花子房、韧皮部汁液和花蜜
中均有分布 ].
2 植物凝集素的定义
植物凝集素 植物凝集素的研究进展
随着植物凝集素分子生物学的研究,对凝集素
蛋白纯化及其结构的研究表明,植物凝集素是一组
高度异质性的具有特异糖结合活性的蛋白.1888年
凝集素被发现时,根据它的特性被命名为血细胞凝
集素(haemagg|utinln),在随后的很长时期内一直
沿用这个名称 直到发现一些植物凝集素选择性地
凝集ABO血型系统中某些类型的血细胞,lectin一
词被提出,用以说明植物凝集素凝集反应的选择性
特征 leetin一词来源于希腊语legere,意思是选
择,但是这个名称并不十分严格,因为它同时包括了
某些具有凝集活性丽非凝集素的蛋白质.从严格意
义上说血细胞凝集素(haemagglutinin)一词更准确
地说明了糖结合蛋白凝集红细胞的能力,但是由于
血细胞凝集素同时具备凝集其他细胞的能力,所以,
凝集素(agglutinin)一词更为台适.尽管haemag一
收藕B期:2001-03-06
作者简介:粱嶂(1966一),男 讲师,现从事强生物学和细胞生物学研究. 维普资讯 http://www.cqvip.com
第2期 粱峰等:植物凝集素的研究进展 233
glutinin、lectin、agglutinin的准确含义并不相同,它
们通常被用以描述同一类蛋白质,在3个名词中,
leetin一词使用相对较多.凝集素最初的定义是指
非免疫来源的糖结台蛋白,可以凝集细胞或沉淀糖
缀合物.最近凝集素的分子生物学研究进展要求根
据凝集素的结构和功能对其进行重新定义.因此,
Peumans和Van Damme在1 995年将植物凝集素
定义为至少具有一个可与单糖或寡聚糖特异可逆结
合的非催化结构域的植物蛋白口].根据这个定义,
一大类具有不同凝集和沉淀糖缀合物能力的植物蛋
白均被包括在凝集素的范畴内.
3 植物凝集素的种类及其糖结合特异性
根据植物凝集素亚基的结构特征,植物凝集素
被分成4种类型:部分凝集素(merolectins)、全凝集
素(hololectins)、嵌台凝集素(chimerolectins)和超
凝集素(superlectins) .根据氨基酸序列的同源
性及其在进化上的关系,植物凝集素可以分为7个
家族:豆科凝集索、几丁质结合凝集素、单子叶甘露
糖结合凝集素、Ⅱ型核糖体失活蛋白、木菠萝素(ja—
ealin)家族、葫芦科韧皮部凝集素和苋科凝集素 ].
本文按后一种分类方法进行介绍.
3.1 豆科凝集素
豆科凝集素是来自豆科植物的一类同源蛋白,
目前已经从7O多种植物中纯化了豆科凝集素,其中
大部分来自种子,豆科凝集素还存在于叶片、茎皮、
植物凝集素 植物凝集素的研究进展
根等营养器官中口’ .目前还发现一些豆科植物具 有两种或多种凝集素,例如荆豆(Ulex europaeus L.)的种子含有两种具有不同糖结合活性的凝集
素,扁豆属(Dotichos biftorus L.)的种子、茎、叶片、 根中至少有4种具有不同糖结合活性的凝集素 ]. 豆科凝集素的糖结合特异性差异很大,大部分属于 甘露糖/葡萄糖和半乳糖/N 乙酰半乳糖胺结台凝 集素,有些识别L岩藻糖、Neu5Aca(2,3)Gal/Gal— NAc,还有的不结台单糖,仅识别寡聚糖.
3.2 几丁质结合凝集隶
许多几丁质结台凝集素均包含有一个或多个橡胶 素(hevein)结构域.橡胶素是从橡胶树的乳液中分离 的、由43个氨基酸组成的小分子质量蛋白质,属于具 有一个几丁质结合位点的单凝集素.目前已经在禾本 科、荨麻科、罂粟科、商陆科和茄科中发现了几丁质结 台凝集素的存在,它们具有高度的序列同渌性。[www.61k.com)].
3.3 单子叶甘露糖结台凝集素
单子叶甘露结台凝集素发现相对较晚.1 987
年,发现了第一个单子叶甘露糖结合凝集素雪花莲 外源凝集素(GNA),它仅具有甘露糖结合专一
性口].之后,结构和进化上相关的植物凝集素相继在 石蒜科、风梨科、天南星科、兰科和百合科中被发现, 都仅对甘露糖具有专一性,对许多单子叶甘露糖结 合凝集素的研究发现,它们属于一个在进化上相关 的家族,不仅序列上具有同源性,而且都表现出对甘 露糖的专一性.单子叶甘露糖结合凝集素由约12 ku的单体组成同源二聚体或四聚体,但是在前体的 加工和修饰上有很大的差别,近年还发现.编码单子 叶甘露糖结合凝集素的基因是一类相关基因组成的 基因家族.对雪花莲属(Galanthus L.)、水仙属等 石蒜科凝集素基因eDNA序列分析表明,它们各自 都有不同的凝集素基因的表达,这可能是有许多同 工凝集素(1ectin isoform)存在的原因[】].
3.4 Ⅱ型核糖体失活蛋白
Ⅱ型核糖体失活蛋白(type 1I ribosome—inacti— vating proteins)具有特异的rRNA N一糖苷酶活性, 能够催化失活真核细胞的核糖体.Ⅱ型核糖体失活 蛋白有1、2或4个相同的亚基组成,每个亚基包括 两个在结构和功能上都不同的多肽.即A链和B
链,两条链通过一个二硫键共价连接.A链具有N一 糖苷酶活性,B链具有糖结合活性.同豆科凝集素 相似,Ⅱ型核糖体失活蛋白也具有不同的糖结合特 异性,主要识别半乳糖或N一乙酰半乳糖胺.目前仅 有少数Ⅱ型核糖体失活蛋白的序列已知.对相思豆
植物凝集素 植物凝集素的研究进展
毒蛋白、蓖麻素、SNA—I和SNA-V(来自Sambu— CUB nigra的茎皮)及SSA(来自Sambucus siebold]一 aria的茎皮)序列的比较,发现Ⅱ型核糖体失活蛋白 序列具有同源性 ].
3.5 术菠萝素(jacalin)家族
木菠萝素是从木菠萝(arotoearpus integrifolia) 的种子中分离的一种可以和半乳糖特异结合的凝集 素.在桑科的其他植物中也发现有类似的凝集素, 以前一直认为桑科凝集素是桑科植物特有的一类小 的凝集素家族,后来人们在其他植物中也发现了与 之序列高度相似的凝集素,于是将它们统归为一个 木菠萝素家族.木菠萝素家族根据其糖结合的专一 性,可以分为两个类别,一类是半乳糖专一性的,包 括木菠萝素和一些桑科凝集素,亚基由一个短的口 链和一个长的a链组成 cJ;另一类是甘露糖专一性 的,都是由多个亚基构成的多聚体,每个亚基由一条 肽链构成0 .
3.6 葫芦科韧皮部凝集索
许多葫芦科植物韧皮部汁液中古有一种二聚体 维普资讯 http://www.cqvip.com
武汉大学学报(理学版) 第48卷
的几丁质结合凝集素,对其序列分析表明,除几丁质 结合特性外,和几丁质结合凝集素并没有序剐上的 同源性,southern杂交分析表明它们由一个小的基 因家族编码口 .
3.7 苋科凝集素
许多苋科植物的种子中包含一类对GaINAc具
有特异性的凝集素,它们在氨基酸序列和三维结构 上同其他的植物凝集素都不同 .
植物凝集素糖结合特异性的另一个重要方面是 它们对外源多糖亲和的优先性.大部分植物凝集素 对动物糖蛋白的亲和性远远高于植物的典型糖类, 还有的凝集素仅仅结合植物不具有或含量很少的糖 类,如几丁质结合凝集素识别真菌细胞壁和无脊椎 动物外骨骼上的多糖,这种对动物和微生物糖类的 优先性表明,植物凝集素的功能可能在于结合外源 糖蛋白而非内源受体.
4 植物凝集素的结构
4.1 豆科凝集素
伴刀豆(Canavalia 玎 rmfs)凝集素(ConA)
等豆科凝集素在结构上非常相似,仅在形成多聚体 的方式和程度上有差异,例如ConA及来自兵豆
(Lens culinaris)、豌豆(Pisum sativum L.)的凝集 素,其单体由两个p片层构成,两个p片层分别由7
植物凝集素 植物凝集素的研究进展
条、6条反向平行的链构成,它们之间通过转折和环 相连,第三个较小的p片层,即s片层将上述两个p 片层连接在一起,S片层由5条较短的链构成(图1, ①)_1 l5].虽然豆科凝集素都具有相似的立体结构, 但组成单体的肽链并不一样,如ConA等Dioclece 族的单链凝集素的前体仅有一条约25 ku的肽链构 成,而~iciece族凝集素的前体由一条2O ku的口链 和一条5 ku的a链通过非共价键结合在一起.
Viciece族凝集素是二聚体,两个单体非共价相连成 一个对称的、十二链的p夹层状结构(图2,①).Co— nA等Dioclece族凝集素依据pH的情况可以形成
二聚体或四聚体(图2,②).上述两种凝集素在糖结 合位点上没有明显的差异.ConA、香豌豆(La—
thyrus odoratus L.)凝集紊(LoL1)和糖的共结晶研 究表明:位于单体上部4个环上的几个氨基酸残基 (LoLl中的Asp 、Oly 、Asn”。(www.61k.com]、Gly跏、A1a加。、 Glu 。)决定了糖结合的特异性,其中有些氨基酸残 基在豆科凝集素中是不变的,如LoLl中的Asp 和 Asn” ,被认为对糖结合活性是必需的,位于一个较 长可变环上的两个残基A1 、GIu 表现出较大的 可变性,被认为可能和糖结合的特异性有关 .
4.2 单子叶甘露糖结合凝集素
在单子叶甘露糖结台凝集素中,对GNA的研究 最为详细.天然GNA是四聚体,有4个12 ku前体 非共价结合成50 ku的同源四聚体.GNA单体肽链
包括109个氨基酸,形成3个亚结构域(I,Ⅱ,III),其 中Ⅱ,Ⅲ 亚结构域均是四链的B片层,I亚结构域是一 个3链的B 片层,它们之间通过环相连
(图1,②)“ ”J.在GNA二聚体中(A和D,B和c), 两条肽链C端的链相互交换,参与到I亚结构域中,使 之也成为一个4链的p片层,这样GNA单体的3个 片层即由12条链组成,3个p片层按棱柱的方式排列 成一个p棱柱状结构,口片层的方向和棱柱的轴垂 直.GNA单体的3个甘露糖结合位点位于有3个0 片层构成的裂隙中,两个GNA二聚体形成GNA四
聚体,形状像一个扁平的王冠,中间有一个较宽的亲 水性通道(图2,③).GNA的糖结合位点有4个保守 氨基酸残基,它们位于I亚结构域中(G1n肼,Asp , Ash9 ,Tyr ),其中Asf As 决定甘舔糖结合特异 性,此外,VaP。也参与了和糖的亲水性结合 .
4.3 术菠萝(jacalin)凝集素
木菠萝素的前体由1 33个氨基酸残基a链和
2O个氨基酸残基B链非共价结合而成,12条链构成 的p片层也排列成p棱柱的形状,B片层的方向同B
植物凝集素 植物凝集素的研究进展
棱柱的轴平行(图1,④) .结台甘磊糖的木菠萝
凝集素,如菊芋(Helianthus tuberosus L.)凝集素
(Hehuba)等,也具有相似的立体结构,仅有一条构
成p片层的链不同(图1,③) .天然的木菠萝素
为四聚体,具有4个Gal/GalNAc结合位点(图1,
⑤).菊芋凝集紊在结晶时形成八聚体,八聚体按圆
环形状排列(图2,④) ,其他结台甘磊糖的木菠萝
凝集紊则形成四聚体或二聚体(图2,⑤)口 .菊芋
凝集素的糖结合位点包括5个氨基酸残基(Oly ,
Oly”。[www.61k.com],Asp ,Val ,Asp 。),它们和甘磊糖通过8
个氢键相连.木菠萝素的糖结合位点和Hehuba的
相似,不同的是其 链N端的Oly 替代了Gly .
4.4 I型核糖体失活蛋白
尽管大部分Ⅱ型RIP和Gal/GalNAc结合(蓖
麻素、相思豆毒蛋白),最近从鸢尾属(Irls L.)植物
Iris hollandica的球茎中分离了两种核糖体失活蛋
白(RIP)IRAr和IRAb可以和半乳糖、甘磊糖结
合口 ,但是IRA B链的结构和其他RIP很相似(图
1,⑤).对IRA B链的结构研究表明,B链的分子质
量为29 ku,包括两个结构域,两个结构域分别包括
4个亚结构域1 、h、1B、17和2 、2a、2B、27.每个
维普资讯 http://www.cqvip.com
第2期 梁峰等:植物凝集素的研究进展
结构域的p片层按三叶草的形式排列,通过a螺旋
相连.两个糖结合位点(Asp 、Se一 、Trp 、
Asn 、G Ln 。)位于p链的两端,即Is和2 亚结构
域上,通过7个氢键和甘露糖结合 .
5 植物凝集素的功能及其应用
现在对植物凝集素的生理作用还有争议,但比
较一致的看法是凝集素的糖结合活性及其专一性决
定了它们的功能.专一性研究表明外源多糖可能是
许多植物凝集素最可能的受体,外源多糖包括真菌
或植物病毒表面、昆虫或草食动物肠道细胞表面的
糖蛋白.基于凝集素和糖的相互作用,对凝集素的
生理作用有许多假说.例如,植物凝集素被认为和
糖的运输.储藏物质的积累、细胞间的互作,以及细
胞分裂的调控等有关.同样,凝集素和外源糖的结
图l 不同类型植物凝集紊前体的三维结构示意图
1.LOLl前体-2 GNA前体的p棱柱结构|3.蒲芋块茎凝集素前体的P棱柱结构-4.术菠萝衰前体的p棱柱结构{5.IRAr的口链构成的三
叶草状结构.★ 表示糖结台位点
维普资讯 http://www.cqvip.com
武汉大学学报(理学版) 第{8卷
合被认为是植物和微生物间的识别、对外源生物防
植物凝集素 植物凝集素的研究进展
御机制建立的决定因素.后来人们认识到植物凝集
素不仅在植物体内起作用,如作为氮源、特异的识别
因子等,而且在植物抵抗外来生物如植物病毒、昆
虫、草食动物的防御反应中也起作用.大多数植物
凝集素存在于储藏器官中,它们既可能作为一种氮
源,也可以在植物受到危害作为一种防御蛋白发挥
功能 .
人工饲养试验表明,许多植物凝集素对咀嚼式
口器昆虫具有毒性,如麦胚凝集素(WGA)、马铃薯
(Solanum tuberosur~t L.)块茎凝集素、花生(Ar —
chishypogaea L.)凝集素、刺苹果种子凝集素 荨麻
属(Urtica L)的凝集素抑制绿豆象幼虫的发
育 ““].最近的工作发现刺吸式口器昆虫如椰粉虱、
图2 不同植物凝集素多聚体的三维结构摸型
1.LOLI对称的二聚体·2.ConA的四集体结构I 3.GNA 由 D和B-c阿十二聚体形成的四聚体结构;4 环状排列的菊芋凝集紊八聚体
结构;5 术菠萝索的四囊体结构.★表示糖结台位点
维普资讯 http://www.cqvip.com
第2期 梁峰荨:植物凝集索的研究进展
叶蝉、蚜虫等对一些单子叶植物甘露糖结凝集素敏
感 ].例如纯化的雪花莲外源凝集素(GNA)和转
gna烟草(Nicotiana tabacum L.)的昆虫饲喂都表
明,GNA可以抗刺吸式口器昆虫,而且可以抗植物
病原性线虫 .上述结果都说明植物凝集素在植物
中具有防御功能.
近年来,植物凝集素在抗虫基因工程,特别是针
对蚜虫等同翅目害虫抗性上的应用受到了愈来愈多
的重视.报道的具有抗蚜活性的基因有植物凝集素
基因、异戊烯基转移酶基因(ipt)和番茄抗线虫基因
(Mi)n 。[www.61k.com] ,其中应用最多的植物凝集素基因是雪花
莲外源凝集素(GNA)基因.报道的转植物凝集素
基因的植物有烟草、水稻、马铃薯、小麦(Triticum
aestivum L.)、番茄(L c0声 rs£cDn esculentum M川)
等,它们表现出了对桃蚜、水稻褐飞虱、菜园夜蛾及
麦长管蚜等害虫的抑制和抗性.例如,Hilder等0
人将gna置于35S启动子的控制下,转化烟草,对转
基因烟草的叶片及整株虫试表明转基因植株对桃蚜
具有抗性,使蚜虫的个体大小显著小于对照,发育受
到影响,生殖力下降.
植物凝集素对昆虫如同翅目昆虫及动物产生毒
害的机理还不清楚,一般认为植物凝集素可能通过
和糖蛋白,如和昆虫围食膜表面、消化道上皮细胞的
糖缀合物、或糖基化的消化酶等结合从而影响昆虫、
动物的对营养吸收,促进消化道中细菌的繁殖和诱
植物凝集素 植物凝集素的研究进展
发病灶,抑制昆虫的生长发育和繁殖,最终达到杀
虫、或对动物产生毒害的作用.
某些浓度很低的凝集素可能参与植物体内或体
外的特定的识别过程,例如豆科植物根部的凝集素
可能参与Rhizobium 和Bradyrhizobium 的识别或
结合,在共生关系的建立中起着决定性园子的作
用 .
近年来的研究还发现,单子叶甘露糖结合凝集
素对人和动物反转录病毒(HIV)具有抑制作用 ,
某些豆科种子凝集素同细菌细胞壁上的胞壁酸、N
乙酰胞壁酸及胞壁二肽有强烈的互作,麦胚凝集素
可以抑制真菌的生长.因此一般认为凝集素可能具
有保护植物抵抗细菌、真菌、植物病毒等植物病原体
侵袭的作用 .
参考文献:
1] Van Damme E J M.Peumans W J.Pusztai A.et a1. Handbook of Plant Lectins:Properties and Biomed— ical Applications[M].Chichester;John Willey and Sons Ltd,l997.
[2] Peumans W J.Van Damme E J M.Lectins as Plant Defense Proteins[J].Plant Physio1.1995,109:347— 352
3] Gao Y.Qu L J.Chen zL Study ontheMolecular Bi o]ogy of Plant Lecdn[J1.Biotechnology Information. 2000,5:18 22.
L4一Peumans W J,Van Damine E J M.Plant Lectins:
Versatile Proteins with Important Perspectives in Bio— technology[J].Biotechnology and Genetic.Engineer— ing Reviews.1998,15:199—228.
[5j Van Damme E J M,Barre A,Smeets K.et al The Bark Lectin of Robinia Pseud0acacia Contains a Com— plex Mixture of Isolectins Characterization of the Pro— reins and the eDNA Clones[J1.Plant Physiol,1995. 107:838—843.
[6] Van Damme E j M,Barre A,Rouge P.et a1.Molu— cular Cloning of the Ba rk and Seed Lectins from the Japanese Pagoda Tree(sophora japonica)[J] Plant M ol Bio1.1997.33:523-536.
[7] Konami Y.Yamamoto K,(1saws T,The Primary Structures of two Types of the Ulex Europeus Seed Lectin[J Jounal of Bioechemistry,1 991,109':550 658.
[82 Etzler M E.Vegetative Tissue Lectins from Doliehos giflorus[J] Methodsin Enzymology,1989,179:341—
植物凝集素 植物凝集素的研究进展
347.
_g] Van Damme E J M,Allen A K Isolation and Charae— terization of a Lectin with Exclusively Specificity To wards Mannose from Snowdrop(Galanthus nivalis)
Bulbs[J~.FEBSLetters,1 987.215:14O一144
[1o]Young N M,johnston R A Z,Watson D C The Ami— mo Acid Sequences of Jacalin and the Maclurapomifera Agglutlnin[j~.FEBSLetter.1991.282:382—384.
[11]Van Damme E J M,Barre A,Mazard A M,et a1. Characterization and Molecular Cloning of the Lectin from Heliznthus Tuherosus[J]. Eur J Biochem,
1999.259:135—142.
[12 Bostwick D E,Skaggs M I.Thompson G A.Organi— zation and Charaeterization of CUCUrbira Phloem LectIn Genes[J].Plant Mol Boil,1994.26:887—897
[13]Raina A,Datta A.Molecular Cloning of a Gene Enco— ding a Seed—specific Protein with Nutritionally Balanced Amino Acid Composition from Amaranthus[J].Proc
Natl Acad Sci USA .1992.89:11774一¨ 778.
[14]Eddman G M.Cunningham B A,Reeke G N.et a1. The Covalent and 1 hrec—dimensionaI Structure of Con canavalin J].Proc Natl Acad Sci USA.1972.69; 2580—2584
[151 Bouckaert J,Hamdrytk T.Wynes L.et a1.Novel 维普资讯 http://www.cqvip.com
238 武汉大学学报(理学版) 第48卷
Stuctures of Plant Lectins and Their Complexes with Carbohydrates[J].Current Opinon in structural Biol— ogy.1999-9:572—577
[18]Boume Y,Roussel A,Prey M,et a1.Three—dimen— s[onaI Structures of Complexes of Lathyrus Ochrus lso]eetin 1 with Glucose and Mannose}Fine Specificity 。(www.61k.com){the Monosaccharidc-binding Site[j Proteins
Struct Funet Genet.1990,8:365—376.
r17] Heater G,Kaku H .Go]dstein 1 J,et Structure of Mammose-spec/fic Sn0wdrop(galanthus nivalis)Lectin is Representative of a New Plant Lecdn Family'[J: Nature Structural Biolagy·1995,2:472—479
[183 Hester G,Wright c S.The Mamraose—specilic Bulb Lect n from Galanthus Nivalis (Snowdrop) Binds
Mono-and DiN2ammosides日t Distinct Sites.Structurc Analysis of Refined Complexes at 2.3A and 3.0A Res— olution[J].J Mog Bio1.1 998,262:516—831.
[193 Sankaranarayanan R,Sekar Kt Bane~ee R,et a1.A
植物凝集素 植物凝集素的研究进展
Novel Mode of Carbohydrate Recognition in Jacalint a Moraceae Plant Lectin with a 8-prism Fotd[1] Nature Struct Bio1.1996.3:596—603.
[20] Bourne Y.Zamboni V.Barre A.et a1.Helianthus Tuberosus Lectin Reveals a Widspread Scaffold /or Mannose—binding Lectins[j].Structure,1999.7:
1478—148Z
[Z1]Hao Q.Van Damme E J Mt Hause B.et aI.Iris(Iris hollandic~,car.Professor Blaauw/Bulbs Express type 1 and type 2 Ribosome-inactivating Proteins with Unu— sual Properties[Jj.Plant Physiol,9001.125:866— 878
[20]Barre A,BourneY,VanDamme E J M,eL a1.Man—
nose-binding Plant Lectirts Different Structural Seed— folds for a Common Sugar—recognition Process[J]. Biochimie,9001.83:845-651
[2s]Murdock L L.Huesing J E,Nielsen S S,eLa1.Btolog— Jcal Effects of Plant Lect[ns on the Cowpea Weevil[J]. Phytochemistry-1990,29:85—89.
[24]Huesing J E,Murdock L L.Shade R E Rice and
Stingil3.g Nettle Lectins:insecticidal Activity Simila~to Wheat Germ Aggtutinin L J一7.Phy~ochemistr>,1991, 30:3565-3569
r25] Hrider V A.Powel】K S,Gatehouse A M R.et al Expression 0f Snowdrop Leetin in Transgenic Tobacco Plants Results in Added Protection Against Aphids
[J].TransgenicResearch,1995,4:18—25.
Ds]Smigocki A,Neat J w,Jr.et a1.Cytoklnin—mediated Insect Resistance in Nitalians Plants Transfromed with the lpt Gene[J].Plant Mol Boil.1993.23:325—335.
[27]Rossi M,Goggin F L,Milligan S B,et a1.The Nema— tode Resistance Gene Mi of Tomato Co nfers Resistance Against the Potato Aphid[J].Pro Nail Acad Sci
A,1998.95 9750—9754.
[28]Diaa C.Melchers L S,Hoeykass P J J.et a1.Root Lectin as a Determiant of Host-plant Specificity in the Rhizobium-legume Symbiosis[J].Nature,I989.338: 579—581.
[893 Mahmood N.Hay A j.An ELISA Utilizing lmmobil— ised Snowdrop Lectin GNA for the Detection of Enve— lope Glycoproteins of H1V and SlY[J2.Journal of
Immunological Methods 1992.151:9-13
[∞]Chrispeels M J,Raikhel N V Lectin~.Lectin Genes. and their Role in Plant Defense~J].Plant Celt.199i.
植物凝集素 植物凝集素的研究进展
3 1—9
Plant Lectin and Its Application in
Insect—Resistant Plant Genetic Engineering
LIANG Feng,CHANG Tuan-jie
(Department of Biology.Shangqiu Teachers Co llege-Shangqiu
476000.HengntChina)
Abstract:Plant lectins are proteins possessing at least one non-catalytic domain that binds reversibly to
specific mono or oIigosaccharides.They distinguish themselves from other plant proteins by the ability of
carbohydrate binding.M ost plant lectins are directed to bind foreign polysacehride.Plant leetin is believed
to take part in the defense responses against invader.In this paper we presented a review on the classifica
tion,characters,functions,crystal structure and functions related to the carbohydrate binding activity.
Key words:plant lectim carbohydrate binding ability;crystal structure;functions; insect—resitant
plant genetic engineering
维普资讯 http://www.cqvip.com
__
本文标题:植物生物技术研究进展-微生物阿魏酸酯酶的研究进展61阅读| 精彩专题| 最新文章| 热门文章| 苏ICP备13036349号-1